
Бактериофаги — это… Что такое Бактериофаги?
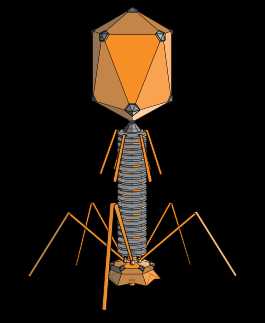
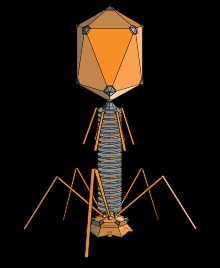 Структура типичного миовируса бактериофага.
Структура типичного миовируса бактериофага.Бактериофа́ги (фаги) (от др.-греч. φᾰγω — «пожираю») — вирусы, избирательно поражающие бактериальные клетки. Чаще всего бактериофаги размножаются внутри бактерий и вызывают их лизис. Как правило, бактериофаг состоит из белковой оболочки и генетического материала одноцепочечной или двуцепочечной нуклеиновой кислоты (ДНК или, реже, РНК). Размер частиц приблизительно от 20 до 200 нм.
История
Английский бактериолог Туорт, Фредерик в статье 1915 года описал инфекционную болезнь стафилококков, инфицирующий агент проходил через фильтры, и его можно было переносить от одной колонии к другой.
Независимо от Фредерика Туорта французско-канадский микробиолог Д’Эрель, Феликс 3 сентября 1917 год сообщил об открытии бактериофагов. Наряду с этим известно, что российский микробиолог Николай Фёдорович Гамалея ещё в 1898 году, впервые наблюдал явление лизиса бактерий (сибиреязвенной палочки) под влиянием перевиваемого агента
После открытия явлений бактериофагии Д’Эрелль развил учение о том, что бактериофаги патогенных бактерий, являясь их паразитами, играют большую роль в патогенезе инфекций, обеспечивая выздоровление больного организма, а затем создания специфического иммунитета. Это положение привлекло к явлению бактериофагии внимание многих исследователей, которые предполагали найти в фагах важное средство борьбы с наиболее опасными инфекционными болезнями человека и животных.
Также Феликс Д’Эрель выдвинул предположение, что бактериофаги имеют корпускулярную природу. Однако только после изобретения электронного микроскопа удалось увидеть и изучить ультраструктуру фагов. Долгое время представления о морфологии и основных особенностях фагов основывались на результатах изучения фагов Т-группы — Т1, Т2,…, Т7, которые размножаются на
Роль бактериофагов в биосфере
Бактериофаги представляют собой наиболее многочисленную, широко распространенную в биосфере и, предположительно, наиболее эволюционно древнюю группу вирусов[3][4]. Приблизительный размер популяции фагов составляет более 1030 фаговых частиц[5].
В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, выделения человека и животных, вода и т. д.) микроорганизмами, тем в большем количестве в нём встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами черноземы и почвы, в которые вносились органические удобрения.
Бактериофаги выполняют важную роль в контроле численности микробных популяций, в автолизе стареющих клеток, в переносе бактериальных генов, выступая в качестве векторных «систем»[6].
Действительно, бактериофаги представляют собой один из основных подвижных генетических элементов. Посредством трансдукции они привносят в бактериальный геном новые гены. Было подсчитано, что за 1 секунду могут быть инфицированы 10
Высокий уровень специализации, долгосрочное существование, способность быстро репродуцироваться в соответствующем хозяине способствует их сохранению в динамичном балансе среди широкого разнообразия видов бактерий в любой природной экосистеме. Когда подходящий хозяин отсутствует, многие фаги могут сохранять способность к инфицированию на протяжении десятилетий, если не будут уничтожены экстремальными веществами либо условиями внешней среды
Строение бактериофагов
1 — головка, 2 — хвост, 3 — нуклеиновая кислота, 4 — капсид, 5 — «воротничок», 6 — белковый чехол хвоста, 7 — фибрилла хвоста, 8 — шипы, 9 — базальная пластинкаБактериофаги различаются по химической структуре, типу нуклеиновой кислоты, морфологии и характеру взаимодействия с бактериями. По размеру бактериальные вирусы в сотни и тысячи раз меньше микробных клеток.
Типичная фаговая частица (вирион) состоит из головки и хвоста. Длина хвоста обычно в 2 — 4 раза больше диаметра головки. В головке содержится генетический материал — одноцепочечная или двуцепочечная РНК или ДНК с ферментом транскриптазой в неактивном состоянии, окруженная белковой или липопротеиновой оболочкой —
Нуклеиновая кислота и капсид вместе составляют нуклеокапсид. Бактериофаги могут иметь икосаэдральный капсид, собранный из множества копий одного или двух специфичных белков. Обычно углы состоят из пентамеров белка, а опора каждой стороны из гексамеров того же или сходного белка. Более того, фаги по форме могут быть сферические, лимоновидные или плеоморфные[10]. Хвост представляет собой белковую трубку — продолжение белковой оболочки головки, в основании хвоста имеется АТФаза, которая регенерирует энергию для инъекции генетического материала. Существуют также бактериофаги с коротким отростком, не имеющие отростка и нитевидные
Фаги, как и все вирусы, являются абсолютными внутриклеточными паразитами. Хотя они переносят всю информацию для запуска собственной репродукции в соответствующем хозяине, у них отсутствуют механизмы для выработки энергии и рибосомы для синтеза белка. У некоторых фагов в геноме содержится несколько тысяч оснований, тогда как фаг G, самый крупный из секвенированных фагов, содержит 480 000 пар оснований — вдвое больше среднего значения для бактерий, хотя всё же недостаточного количества генов для важнейшего бактериального органоида как рибосомы.
Систематика бактериофагов
Большое количество выделенных и изученных бактериофагов определяет необходимость их систематизации. Классификация вирусов бактерий претерпевала изменения: основывалась на характеристике хозяина вируса, учитывались серологические, морфологические свойства, а затем строение и физико-химический состав вириона
В настоящее время согласно Международной классификации и номенклатуре вирусов бактериофаги, в зависимости от типа нуклеиновой кислоты разделяют на ДНК- и РНК- содержащие.
По морфологическим характеристикам ДНК-содержащие фаги выделены в следующие семейства: Myoviridae, Siphoviridae, Podoviridae, Lipothrixviridae, Plasmaviridae, Corticoviridae, Fuselloviridae, Tectiviridae, Microviridae, Inoviridae Plectovirus и Inoviridae Inovirus.
РНК-содержащие: Cystoviridae, Leviviridae[13].
Взаимодействие бактериофага с бактериальными клетками
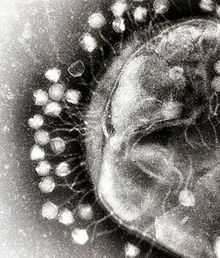 Адсорбция бактериофагов на поверхности бактериальной клетки
Адсорбция бактериофагов на поверхности бактериальной клеткиПо характеру взаимодействия бактериофага с бактериальной клеткой различают вирулентные и умеренные фаги[11]. Вирулентные фаги могут только увеличиваться в количестве посредством литического цикла[8]. Процесс взаимодействия вирулентного бактериофага с клеткой складывается из нескольких стадий: адсорбции бактериофага на клетке, проникновения в клетку, биосинтеза компонентов фага и их сборки, выхода бактериофагов из клетки[7][14].
Первоначально бактериофаги прикрепляются к фагоспецифическим рецепторам на поверхности бактериальной клетки. Хвост фага с помощью ферментов, находящихся на его конце (в основном лизоцима), локально растворяет оболочку клетки, сокращается и содержащаяся в головке ДНК инъецируется в клетку, при этом белковая оболочка бактериофага остается снаружи. Инъецированная ДНК вызывает полную перестройку метаболизма клетки: прекращается синтез бактериальной ДНК, РНК и белков. ДНК бактериофага начинает транскрибироваться с помощью собственного фермента транскриптазы, который после попадания в бактериальную клетку активируется. Синтезируются сначала ранние, а затем поздние иРНК, которые поступают на рибосомы клетки-хозяина, где синтезируются ранние (ДНК-полимеразы, нуклеазы) и поздние (белки капсида и хвостового отростка, ферменты лизоцим, АТФаза и транскриптаза) белки бактериофага. Репликация ДНК бактериофага происходит по полуконсервативному механизму и осуществляется с участием собственных ДНК-полимераз. После синтеза поздних белков и завершения репликации ДНК наступает заключительный процесс — созревание фаговых частиц или соединение фаговой ДНК с белком оболочки и образование зрелых инфекционных фаговых частиц
Продолжительность этого процесса может составлять от нескольких минут до нескольких часов[8]. Затем происходит лизис клетки, и освобождаются новые зрелые бактериофаги[11]. Иногда фаг инициирует лизирующий цикл, что приводит к лизису клетки и освобождению новых фагов. В качестве альтернативы фаг может инициировать лизогенный цикл, при котором он вместо репликации обратимо взаимодействует с генетической системой клетки-хозяина, интегрируясь в хромосому или сохраняясь в виде плазмиды
Очень важным свойством бактериофагов является их специфичность: бактериофаги лизируют культуры определенного вида, более того, существуют так называемые типовые бактериофаги, лизирующие варианты внутри вида, хотя встречаются поливалентные бактериофаги, которые паразитируют в бактериях разных видов[16][17].
Жизненный цикл
Умеренные и вирулентные бактериофаги на начальных этапах взаимодействия с бактериальной клеткой имеют одинаковый цикл.
- Адсорбция бактериофага на фагоспецифических рецепторах клетки.
- Инъекция фаговой нуклеиновой кислоты в клетку хозяина.
- Совместная репликация фаговой и бактериальной нуклеиновой кислоты.
- Деление клетки.
- Далее бактериофаг может развиваться по двум моделям: лизогенный либо литический путь.
- Умеренные бактериофаги после деления клетки находятся в состоянии профага (Лизогенный путь).
- Вирулентные бактериофаги развиваются по Литической модели:
- Нуклеиновая кислота фага направляет синтез ферментов фага, используя для этого белоксинтезирующий аппарат бактерии. Фаг тем или иным способом инактивирует ДНК и РНК хозяина, а ферменты фага совсем расщепляют её; РНК фага «подчиняет» себе клеточный аппарат синтеза белка.
- Нуклеиновая кислота фага реплицируется, и направляет синтез новых белков оболочки. Образуются новые частицы фага в результате спонтанной самосборки белковой оболочки (капсид) вокруг фаговой нуклеиновой кислоты; под контролем РНК фага синтезируется лизоцим.
- Лизис клетки: клетка лопается под воздействием лизоцима; высвобождается около 200—1000 новых фагов; фаги инфицируют другие бактерии.
Применение
В медицине
Одной из областей использования бактериофагов является антибактериальная терапия, альтернативная приёму антибиотиков. Например, применяются бактериофаги: стрептококковый, стафилококковый, клебсиеллёзный, дизентерийный поливалентный, пиобактериофаг, коли, протейный и колипротейный и другие.
Бактериофаги применяются также в генной инженерии в качестве векторов, переносящих участки ДНК, возможна также естественная передача генов между бактериями посредством некоторых фагов (трансдукция).
Фаговые векторы обычно создают на базе умеренного бактериофага λ, содержащего двухцепочечную линейную молеклул ДНК. Левое и правое плечи фага имеют все гены, необходимые для литического цикла (репликации, размножения). Средняя часть генома бактериофага λ (содержит гены, контролирующие лизогению, то есть его интеграцию в ДНК бактериальной клетки) не существенна для его размножения и составляет примерно 25 тысяч пар нуклеотидов. Данная часть может быть заменена на чужеродный фрагмент ДНК. Такие модифицированные фаги проходят литический цикл, но лизогения не происходит. Векторы на основе бактериофага λ используют для клонирования фрагментов ДНК эукариот (то есть более крупных генов) размером до 23 т.п.н. Причем, фаги без вставок — менее 38 т.п.н или, напротив, со слишком большими вставками — более 52 т.п.н не развиваются и не поражают бактерии[18].
В биологии
Бактериофаги M13, фаг Т4, T7 и фаг λ используют для изучения белок-белковых, белок-пептидных и ДНК-белковых взаимодействий методом фагового дисплея.
Поскольку размножение бактериофага возможно только в живых клетках бактериофаги могут быть использованы для определения жизнеспособности бактерий. Данное направление имеет большие перспективы, поскольку, одним из основных вопросов при разных биотехнологических процессах является определение жизнеспособности используемых культур. С помощью метода электрооптического анализа клеточных суспензий была показана возможность изучения этапов взаимодействия фаг-микробная клетка[19].
Ссылки
- ↑ Вирусы бактерий
- ↑ Бактериофаг
- ↑ Ackermann H.-W. // Res. Microbiol., 2003. — V. 154. — P. 245—251
- ↑ Hendrix R.W. // Theor. Popul. Biol., 2002. — V. 61. — P. 471—480
- ↑ Suttle C.A. (September 2005), Vuiruses in the sea. Nature 437:356-361.
- ↑ Шестаков С. В. Как происходит и чем лимитируется горизонтальный перенос генов у бактерий. Экологическая генетика 2007. — Т. 5. — № 2. — C. 12-24.
- ↑ 1 2 Tettelin H., Masignani V., Cieslewicz M. J., Donati C., Medini D., Ward N. L., Angiuoli S. V., Crabtree J., Jones A. L., Durkin A. S., Deboy R. T., Davidsen T. M., Mora M., Scarselli M., Margarit y Ros I., Peterson J. D., Hauser C. R., Sundaram J. P., Nelson W. C., Madupu R., Brinkac L. M., Dodson R. J., Rosovitz M. J., Sullivan S. A., Daugherty S. C., Haft D. H., Selengut J., Gwinn M. L., Zhou L., Zafar N., Khouri H., Radune D., Dimitrov G., Watkins K., O’Connor K. J., Smith S., Utterback T. R., White O., Rubens C. E., Grandi G., Madoff L. C., Kasper D. L., Telford J. L.,. Wessels M. R, Rappuoli R., Fraser C. M. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: implications for the microbial «pan-genome.» Proc. Natl. Acad. Sci. USA 2005. 102: 13950-13955
- ↑ 1 2 3 4 5 Guttman B., Raya R., Kutter E. Basic Phage Biology, in Bacteriophages: Biology and Applications, (Kutter E. and Sulakvelidze A., ed.), CRP Press, 2005 FL. — Р.29-66.
- ↑ Ковалева Е. Н. Создание биопрепарата на основе выделенных и изученных бактериофагов Enterococcus faecalis: Дис. … канд. биол. наук. — Саратов, 2009. — 151 с
- ↑ Ackermann H.-W. // Res. Microbiol., 2003. — V. 154. — P. 245—251.
- ↑ 1 2 3 4 Ожерельева Н. Г. Краткая Медицинская Энциклопедия, М.: изд-во «Советская Энциклопедия», 1989. — издание второе.
- ↑ Русалеев В. С., Таксономия вирусов бактерий / В. С. Русалеев // Ветеринария. — 1990. — № 12. — C. 25-28.
- ↑ Virus Taxonomy. Classification and Nomenclature of Viruses. Seventh Report of the International Committee on Taxonomy of Viruses / Edited by M.H.V. van Regenmontel et al. — San Diego: Academic Press, 2000. — P. 43-53, 64-129.
- ↑ Raya R.R., Hébert E.M. Isolation of phage via induction of lysogens. Bacteriophages: Methods and Protocols, Volume 1: Isolation, Characterization, and Interaction (Martha R.J. Clokie, Andrew M. Kropinski (eds.), 2009. — V. 501. — P. 23-32.
- ↑ 1 2 Микробиология: учеб. пособие / В. В. Лысак. — Минск : БГУ, 2007. — 430 с.
- ↑ Адамс М., Бактериофаги / М. Адамс. — М.:Медгиз, 1961. — 521 с.
- ↑ Гольдфарб Д. М., Бактериофагия / Д. М. Гольдфарб. — М.: Медгиз, 1961. — 299 с.
- ↑ Щелкунов С. Н. Генетическая инженерия / С. Н. Щелкунов. — Новосибирск : Сиб. унив. изд-во, 2004. — 496 с.
- ↑ Guliy O.I., Bunin V.D., O’Neil D., Ivnitski D., Ignatov O.V. A new electro-optical approach to rapid assay of cell viability // Biosensors and Bioelectronics. 2007. V. 23. P. 583—587.
dic.academic.ru
Бактериофаги виды и назначение, список препаратов, применение бактериофагов в медицине, лечение комплексными бактериофагами, классификация и строение
Бактериофагами являются специфические вирусы, избирательно атакующие и поражающие микробы. Размножаясь внутри клетки, они разрушают бактерии. При этом патогенная микрофлора уничтожается, а полезная – сохраняется.
Использование этих вирусов было предложено еще в начале века для лечения инфекционных заболеваний. Однако интерес к ним во многих странах мира был утрачен после появления антибиотиков. Сегодня интерес к данным вирусам возвращается.
Вконтакте
Google+
Мой мир
Особенности строения и среда обитания
Что такое бактериофаги ? Это большая группа вирусов, размером в 100 раз меньше клеток бактерий. Структура фагов при многократном увеличении поражает многообразием.
Какие бывают бактериофаги
 Рассмотрим виды микробов и назначение, в зависимости от их типа.
Рассмотрим виды микробов и назначение, в зависимости от их типа.
Существует девятнадцать семейств вирусов, различающихся по типу нуклеиновой кислоты (ДНК или РНК), а также по форме и строению генома.
Бактериофаги в медицине классифицируются в соответствии со скоростью влияния на патогенной бактерии:
- Умеренные бактериофаги медленно и частично разрушают болезнетворные микроорганизмы, вызывая в них необратимые изменения, передающиеся следующему поколению микробов. Это так называемый лизогенный эффект.
- Вирулентные молекулы вируса, попав в клетки микроба, активно и быстро размножаются. Они приводят к гибели бактерии почти мгновенно (литический эффект).
- Умеренный вид микробов применяют как альтернативу для лечения бактериальных инфекций. Они имеют определенные преимущества:
- Удобная форма. Препарат производится для перорального приема как раствор или в виде таблетки.
В отличие от антибиотиков, бактериофаги не имеют побочных действий, они реже вызывают аллергическую реакцию, не имеют вторичных негативных эффектов.
Отсутствует резистентность микробов. Бактериям адаптироваться к вирусам сложнее, а при комплексном воздействии это почти невозможно.
Но есть и свои минусы:
- курс терапии более длительный;
- определенные трудности с выбором правильной группы препаратов;
- геном бактерии переносится от одного микроба к другому.
Виды бактериофагов
В медицине, с учетом специфичности описываемых вирусов, предпочитают применять комплексные и поливалентные бактериофаги, которые содержат несколько разновидностей этих микробов.
Список и описание бактериофагов:
- Дизфак, поливалентный дизентерийный. Он вызывает гибель шигелл Флекснера и Зонне.
- Брюшнотифозный убивает возбудителей брюшного тифа, сальмонелл.
- Клебсиеллезный поливалентный. Представляет собой комплексное средство, уничтожающее клебсиеллы пневмонии, озена, риносклеромы.
- Клебсиелл пневмонии, клебсифаг – отличный помощник в борьбе с урогенитальной, дыхательной, пищеварительной систем, хирургических инфекций, генерализованных септических патологий.
- Колипротеофаг, колипротейный. Предназначается для лечения пиелонефритов, циститов, колитов и других болезней, спровоцированных протеем и кишечными палочками.
- Колифаг, коли. Эффективно действует в терапии инфекций кожи и внутренних органов, спровоцированной энтеропатогенной кишечной палочкой Е. Coli.
- Протеофаг, протейный губительно действует на специфические протейные микробы вульгарис и мирабилис, являющиеся возбудителями гнойных воспалений патологий кишечника.
- Стрептококковы, стрептофаг быстро нейтрализует стафилококки, выделяемые при любых гнойных инфекциях.
- Синегнойный. Рекомендуется для терапии воспалений, который провоцирует синегнойная палочка. Лизирует бактерии псевдомонас аэругиноза.
- Комплексный пиобактериофаг. Является смесью фаголизатов стрептококков, энтерококков, стафилококков, псевдоманус аэругиноза, эшерихий коли, клебсиелл окситока и пневмонии.
- Сектафаг, поливалетный пиобактериофаг. Пагубно действует на эшерихии коли .
- Интенси. Комплексный препарат, лизирующий шигиллы, сальмонеллы, энерококки, стафилококки, псевдоманис протей и аэрунина.
Лечение бактериофагами
 Только врач после обследования и выявления инфекции должен назначать медикаменты. Самостоятельное их применение может быть неэффективным потому, что нельзя определить чувствительность к фагам без специального исследования.
Только врач после обследования и выявления инфекции должен назначать медикаменты. Самостоятельное их применение может быть неэффективным потому, что нельзя определить чувствительность к фагам без специального исследования.
Схема лечения разрабатывается индивидуально для каждого клиента. Чаще всего прибегают к медикаментам для терапии кишечного дисбактериоза. Курс лечения может составлять около пяти дней, но в некоторых случаях – до 15 дней. Повторяют курсы для большей эффективности 2-3 раза.
Пример курса терапии стафилококковой инфекции:
- ребенку до полугода – по 5 мл;
- от полугода до одного года – по 10 мл;
- ребенку от одного года до трех лет – 15 мл;
- от 3-х лет до 8-20 мл;
- ребенку после восьми лет – 30мл.;
- грудничкам дают фаги перорально, каплями в нос, в форме клизмы.
Препараты бактериофагов
 Бактериофаги размножаются внутри бактерий, этим убивая их. В то время как лекарства в процессе лечения расходуются и их количество уменьшается, то количество фагов может, наоборот, возрастать.
Бактериофаги размножаются внутри бактерий, этим убивая их. В то время как лекарства в процессе лечения расходуются и их количество уменьшается, то количество фагов может, наоборот, возрастать.
При исчезновении пищи фагов — вредных бактерий, исчезают и сами фаги.
Препараты бактериофагов применяют при лечении заболеваний у детей:
- ушные инфекции;
- инфекции мочеполовых органов;
- инфекции органов дыхания;
- хирургические инфекции;
- инфекции желудочно-кишечного тракта;
- инфекции глаз и др.
Чтобы вырастить бактериофаги, на питательную среду, которая засеяна определенной культурой бактерий, наносится материал с бактериофагами. В местах их попадания формируется зона разрушенных бактерий, представляющая собой пустое пятно. Данный материал бактериологичной иглой забирается. Его переносят в содержащую бактериальную молодую культуру, суспензию. Эти действия выполняют до 10 раз, чтобы полученная бактериофага была чистой.
Бактериофаги – альтернатива антибиотикам
На основе бактериофагов выпускают препараты в виде свечей, аэрозолей, таблеток, растворов и других форм. В названии медикаментов используют группу бактерий, для борьбы с которыми они предназначается .
Сравнение с антибиотиками
 В отличие от антибиотиков, все виды бактериофагов препаратов не влияют отрицательно на организм человека.
В отличие от антибиотиков, все виды бактериофагов препаратов не влияют отрицательно на организм человека.
Каждый вид избирательно влияет на микроорганизмы, поэтому они не только не вредят микрофлоре, но и применяются при лечении дисбактериоза. Однако эти лекарства используют гораздо реже, чем антибиотики по нескольким причинам:
- Бактериофаги не проникают в кровь. Их применяют только при наличии возможности легко доставить препарат к месту воздействия. Например, прополоскать горло, нанести непосредственно на рану, выпить при кишечной инфекции.
- Для применения бактериофагов важно быть уверенным в диагнозе. Исключением являются комбинированные препараты с бактериофагами против различных возбудителей. Эффективность этих препаратов меньше, а цена больше.
Вконтакте
Google+
Мой мир
lekarstva.guru
Бактериофаги — Википедия
| Группа вирусов | |
 Структура типичного миовируса бактериофага. Структура типичного миовируса бактериофага. | |
| Название | |
|---|---|
| Бактериофаги | |
| Статус названия | |
| не определён | |
| Родительский таксон | |
| Домен Вирусы | |
| Представители | |
Все вирусы, поражающие бактерий | |
Бактериофа́ги или фа́ги (от др.-греч. φᾰγω «пожираю») — вирусы, избирательно поражающие бактериальные клетки. Чаще всего бактериофаги размножаются внутри бактерий и вызывают их лизис. Как правило, бактериофаг состоит из белковой оболочки и генетического материала одноцепочечной или двуцепочечной нуклеиновой кислоты (ДНК или, реже, РНК). Общая численность бактериофагов в природе примерно равна общей численности бактерий (1030{\displaystyle 10^{30}} — 1032{\displaystyle 10^{32}} частиц)[1]. Бактериофаги активно участвуют в круговороте химических веществ и энергии, оказывают заметное влияние на эволюцию микробов и бактерий[1].
История
Английский бактериолог Фредерик Туорт в статье 1915 года описал инфекционную болезнь стафилококков, возбудитель которой проходил через фильтры, и его можно было переносить от одной колонии к другой.
Независимо от Фредерика Туорта французско-канадский микробиолог Феликс Д’Эрелль 3 сентября 1917 года сообщил об открытии бактериофагов.[2] Наряду с этим известно, что российский микробиолог Николай Фёдорович Гамалея ещё в 1897 году впервые наблюдал явление лизиса бактерий (сибиреязвенной палочки) под влиянием перевиваемого агента[3][4].
После открытия явлений бактериофагии Д’Эрелль развил учение о том, что бактериофаги патогенных бактерий, являясь их паразитами, играют большую роль в патогенезе инфекций, обеспечивая выздоровление больного организма, а затем создания специфического иммунитета. Это положение привлекло к явлению бактериофагии внимание многих исследователей, которые предполагали найти в фагах важное средство борьбы с наиболее опасными инфекционными болезнями человека и животных.
Также Феликс Д’Эрелль выдвинул предположение, что бактериофаги имеют корпускулярную природу. Однако только после изобретения электронного микроскопа удалось увидеть и изучить ультраструктуру фагов. Долгое время представления о морфологии и основных особенностях фагов основывались на результатах изучения фагов группы Т — Т1, Т2,…, Т7, которые размножаются на Е. coli штамма B. Однако с каждым годом появлялись новые данные, касающиеся морфологии и структуры разнообразных фагов, что обусловило необходимость их морфологической классификации.
Роль бактериофагов в биосфере
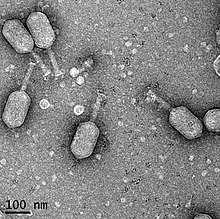 Бактериофаг ϕpp2 патогенных вибрионов V. parahaemolyticus
Бактериофаг ϕpp2 патогенных вибрионов V. parahaemolyticusи V. alginolyticus
Бактериофаги представляют собой наиболее многочисленную, широко распространённую в биосфере и, предположительно, наиболее эволюционно древнюю группу вирусов[5][6]. Приблизительный размер популяции фагов составляет более 1030 фаговых частиц[7].
В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, выделения человека и животных, вода и т. д.) микроорганизмами, тем в большем количестве в нём встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами чернозёмы и почвы, в которые вносились органические удобрения.
Бактериофаги выполняют важную роль в контроле численности микробных популяций, в автолизе стареющих клеток, в переносе бактериальных генов, выступая в качестве векторных «систем»[8].
Действительно, бактериофаги представляют собой один из основных подвижных генетических элементов. Посредством трансдукции они привносят в бактериальный геном новые гены. Было подсчитано, что за 1 секунду могут быть инфицированы 1024 бактерий[9]. Это означает, что постоянный перенос генетического материала распределяется между бактериями, обитающими в сходных условиях.
Высокий уровень специализации, долгосрочное существование, способность быстро репродуцироваться в соответствующем хозяине способствует их сохранению в динамичном балансе среди широкого разнообразия видов бактерий в любой природной экосистеме. Когда подходящий хозяин отсутствует, многие фаги могут сохранять способность к инфицированию на протяжении десятилетий, если не будут уничтожены экстремальными веществами либо условиями внешней среды[10].
Строение бактериофагов
1 — головка, 2 — хвост, 3 — нуклеиновая кислота, 4 — капсид, 5 — «воротничок», 6 — белковый чехол хвоста, 7 — фибрилла хвоста, 8 — шипы, 9 — базальная пластинкаБактериофаги различаются по химической структуре, типу нуклеиновой кислоты, морфологии и характеру взаимодействия с бактериями. По размеру бактериальные вирусы в сотни и тысячи раз меньше микробных клеток.
Типичная фаговая частица (вирион) состоит из головки и хвоста. Длина хвоста обычно в 2—4 раза больше диаметра головки. В головке содержится генетический материал — одноцепочечная или двуцепочечная РНК или ДНК с ферментом транскриптазой в неактивном состоянии, окружённая белковой или липопротеиновой оболочкой — капсидом, сохраняющим геном вне клетки[11].
Нуклеиновая кислота и капсид вместе составляют нуклеокапсид. Бактериофаги могут иметь икосаэдральный капсид, собранный из множества копий одного или двух специфичных белков. Обычно углы состоят из пентамеров белка, а опора каждой стороны из гексамеров того же или сходного белка. Более того, фаги по форме могут быть сферические, лимоновидные или плеоморфные[12].
Хвост, или отросток, представляет собой белковую трубку — продолжение белковой оболочки головки, в основании хвоста имеется АТФаза, которая регенерирует энергию для инъекции генетического материала. Существуют также бактериофаги с коротким отростком, не имеющие отростка и нитевидные[13].
Головка округлой, гексагональной или палочковидной формы диаметром 45—140 нм. Отросток толщиной 10—40 и длиной 100—200 нм. Одни из бактериофагов округлы, другие нитевидны, размером 8×800 нм. Длина нити нуклеиновой кислоты во много раз превышает размер головки, в которой находится в скрученном состоянии, и достигает 60—70 мкм. Отросток имеет вид полой трубки, окружённой чехлом, содержащим сократительные белки, подобные мышечным. У ряда вирусов чехол способен сокращаться, обнажая часть стержня. На конце отростка у многих бактериофагов имеется базальная пластинка, от которой отходят тонкие длинные нити, способствующие прикреплению фага к бактерии. Общее количество белка в частице фага — 50-60%, нуклеиновых кислот — 40-50%.[14]
Фаги, как и все вирусы, являются абсолютными внутриклеточными паразитами. Хотя они содержат всю информацию для запуска собственной репродукции в соответствующем хозяине, у них нет механизмов для выработки энергии и рибосом для синтеза белка. Размер известных фаговых геномов варьирует от нескольких тысяч до 498 тысяч пар оснований (геном фага G, поражающего бацилл)[15][16].
Систематика бактериофагов
Большое количество выделенных и изученных бактериофагов определяет необходимость их систематизации. Этим занимается Международный комитет по таксономии вирусов (ICTV). В настоящее время, согласно Международной классификации и номенклатуре вирусов, бактериофаги разделяют в зависимости от типа нуклеиновой кислоты и морфологии.
На данный момент выделяют девятнадцать семейств. Из них только два РНК-содержащих и только пять семейств имеют оболочку. Из семейств ДНК-содержащих вирусов только два семейства имеют одноцепочечные геномы. У девяти ДНК-содержащих семейств геном представлен кольцевой ДНК, а у других девяти — линейной. Девять семейств специфичны только для бактерий, остальные девять только для архей, а (Tectiviridae) инфицирует как бактерий, так и архей[17].
| Порядок | Семейство | Морфология | Нуклеиновая кислота | Пример |
|---|---|---|---|---|
| Caudovirales | Myoviridae | Без оболочки, сократительный хвост | Линейная дцДНК | Фаг Т4, фаг μ, PBSX, P1Puna-like, P2, I3, Bcep 1, Bcep 43, Bcep 78 |
| Siphoviridae | Без оболочки, несократительный хвост (длинный) | Линейная дцДНК | Фаг λ, фаг T5, phi, C2, L5, HK97, N15 | |
| Podoviridae | Без оболочки, несократительный хвост (короткий) | Линейная дцДНК | Фаг T7, фаг T3, P22, P37 | |
| Ligamenvirales | Lipothrixviridae | В оболочке, палочкообразные | Линейная дцДНК | Вирус »Acidianus filamentous» 1 |
| Rudiviridae | Без оболочки, палочкообразные | Линейная дцДНК | Палочкообразный вирус »Sulfolobus islandicus» 1 | |
| Неизвестен | Ampullaviridae | В оболочке, бутылкообразные | Линейная дцДНК | |
| Bicaudaviridae | Без оболочки, лемонообразные | Кольцевая дцДНК | ||
| Clavaviridae | Без оболочки, палочкообразные | Кольцевая дцДНК | ||
| Corticoviridae | Без оболочки, изометрические | Кольцевая дцДНК | ||
| Cystoviridae | В оболочке, сферические | Сегментированная дцРНК | ||
| Fuselloviridae | Без оболочки, лемонообразные | Кольцевая дцДНК | ||
| Globuloviridae | В оболочке, изометрические | Линейная дцДНК | ||
| Guttaviridae | Без оболочки, яйцевидные | Кольцевая дцДНК | ||
| Inoviridae | Без оболочки, нитевидные | Кольцевая оцДНК | M13 | |
| Leviviridae | Без оболочки, изометрические | Линейная оцРНК | MS2, Qβ | |
| Microviridae | Без оболочки, изометрические | Кольцевая оцДНК | ΦX174 | |
| Plasmaviridae | В оболочке, плеоморфные | Кольцевая дцДНК | ||
| Tectiviridae | Без оболочки, изометрические | Линейная дцДНК |
Взаимодействие бактериофага с бактериальными клетками
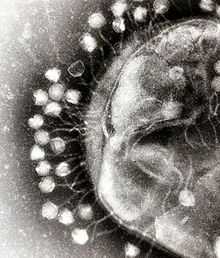 Адсорбция бактериофагов на поверхности бактериальной клетки
Адсорбция бактериофагов на поверхности бактериальной клеткиПо характеру взаимодействия бактериофага с бактериальной клеткой различают вирулентные и умеренные фаги[13]. Вирулентные фаги могут только увеличиваться в количестве посредством литического цикла[10]. Процесс взаимодействия вирулентного бактериофага с клеткой складывается из нескольких стадий: адсорбции бактериофага на клетке, проникновения в клетку, биосинтеза компонентов фага и их сборки, выхода бактериофагов из клетки[9][19].
Первоначально бактериофаги прикрепляются к фагоспецифическим рецепторам на поверхности бактериальной клетки. Хвост фага с помощью ферментов, находящихся на его конце (в основном лизоцима), локально растворяет оболочку клетки, сокращается и содержащаяся в головке ДНК инъецируется в клетку, при этом белковая оболочка бактериофага остаётся снаружи. Инъецированная ДНК вызывает полную перестройку метаболизма клетки: прекращается синтез бактериальной ДНК, РНК и белков. ДНК бактериофага начинает транскрибироваться с помощью собственного фермента транскриптазы, который после попадания в бактериальную клетку активируется. Синтезируются сначала ранние, а затем поздние иРНК, которые поступают на рибосомы клетки-хозяина, где синтезируются ранние (ДНК-полимеразы, нуклеазы) и поздние (белки капсида и хвостового отростка, ферменты лизоцим, АТФаза и транскриптаза) белки бактериофага. Репликация ДНК бактериофага происходит по полуконсервативному механизму и осуществляется с участием собственных ДНК-полимераз. После синтеза поздних белков и завершения репликации ДНК наступает заключительный процесс — созревание фаговых частиц или соединение фаговой ДНК с белком оболочки и образование зрелых инфекционных фаговых частиц[20].
Продолжительность этого процесса может составлять от нескольких минут до нескольких часов[10]. Затем происходит лизис клетки, и освобождаются новые зрелые бактериофаги[13]. Иногда фаг инициирует лизирующий цикл, что приводит к лизису клетки и освобождению новых фагов. В качестве альтернативы фаг может инициировать лизогенный цикл, при котором он вместо репликации обратимо взаимодействует с генетической системой клетки-хозяина, интегрируясь в хромосому или сохраняясь в виде плазмиды[10]. Таким образом, вирусный геном реплицируется синхронно с ДНК хозяина и делением клетки, а подобное состояние фага называется профагом. Бактерия, содержащая профаг, становится лизогенной до тех пор, пока при определённых условиях или спонтанно профаг не будет стимулирован на осуществление лизирующего цикла репликации. Переход от лизогении к лизису называется лизогенной индукцией или индукцией профага. На индукцию фага оказывает сильное воздействие состояние клетки хозяина предшествующее индукции, также как наличие питательных веществ и другие условия, имеющие место в момент индукции. Скудные условия для роста способствуют лизогенному пути, тогда как хорошие условия способствуют лизирующей реакции[10][13][20].
Очень важным свойством бактериофагов является их специфичность: бактериофаги лизируют культуры определённого вида, более того, существуют так называемые типовые бактериофаги, лизирующие варианты внутри вида, хотя встречаются поливалентные бактериофаги, которые паразитируют в бактериях разных видов[21][22].
Жизненный цикл
Умеренные и вирулентные бактериофаги на начальных этапах взаимодействия с бактериальной клеткой имеют одинаковый цикл.
- Адсорбция бактериофага на фагоспецифических рецепторах клетки.
- Инъекция фаговой нуклеиновой кислоты в клетку хозяина.
- Совместная репликация фаговой и бактериальной нуклеиновой кислоты.
- Деление клетки.
- Далее бактериофаг может развиваться по двум моделям: лизогенный либо литический путь.
- Умеренные бактериофаги после деления клетки находятся в состоянии профага (лизогенный путь).
- Вирулентные бактериофаги развиваются по литической модели:
- Нуклеиновая кислота фага направляет синтез ферментов фага, используя для этого белоксинтезирующий аппарат бактерии. Фаг тем или иным способом инактивирует ДНК и РНК хозяина, а ферменты фага совсем расщепляют её; РНК фага «подчиняет» себе клеточный аппарат синтеза белка.
- Нуклеиновая кислота фага реплицируется и направляет синтез новых белков оболочки. Образуются новые частицы фага в результате спонтанной самосборки белковой оболочки (капсид) вокруг фаговой нуклеиновой кислоты; под контролем РНК фага синтезируется лизоцим.
- Лизис клетки: клетка лопается под воздействием лизоцима; высвобождается около 200—1000 новых фагов; фаги инфицируют другие бактерии.
Применение
В медицине
Одной из областей использования бактериофагов является антибактериальная терапия, альтернативная приёму антибиотиков. Например, применяются бактериофаги: стрептококковый, стафилококковый, клебсиеллёзный, дизентерийный поливалентный, пиобактериофаг, коли, протейный и колипротейный и другие. В России зарегистрировано и применяется 13 медицинских препаратов на основе фагов[1]. В настоящее время их применяют для лечения бактериальных инфекций, которые не чувствительны к традиционному лечению антибиотиками, особенно в республике Грузия[23][24][25]. Обычно, применение бактериофагов сопровождается большим, чем антибиотики, успехом там, где присутствуют биологические мембраны, покрытые полисахаридами, через которые антибиотики обычно не проникают[26]. В настоящее время терапевтическое применение бактериофагов не получило одобрения на Западе, хотя и применяются фаги для уничтожения бактерий, вызывающих пищевые отравления, таких, как листерии[27]. В многолетнем опыте в объёме крупного города и сельской местности доказана необычайно высокая лечебная и профилактическая эффективность дизентерийного бактериофага (П. М. Лернер, 2010)[источник не указан 1950 дней]. В России терапевтические фаговые препараты делают давно, фагами лечили ещё до антибиотиков. В последние годы фаги широко использовали после наводнений в Крымске[28] и Хабаровске, чтобы предотвратить дизентерию.
В биологии
Бактериофаги применяются в генной инженерии в качестве векторов, переносящих участки ДНК, возможна также естественная передача генов между бактериями посредством некоторых фагов (трансдукция).
Фаговые векторы обычно создают на базе умеренного бактериофага λ, содержащего двухцепочечную линейную молекулу ДНК. Левое и правое плечи фага имеют все гены, необходимые для литического цикла (репликации, размножения). Средняя часть генома бактериофага λ (содержит гены, контролирующие лизогению, то есть его интеграцию в ДНК бактериальной клетки) не существенна для его размножения и составляет примерно 25 тысяч пар нуклеотидов. Данная часть может быть заменена на чужеродный фрагмент ДНК. Такие модифицированные фаги проходят литический цикл, но лизогения не происходит. Векторы на основе бактериофага λ используют для клонирования фрагментов ДНК эукариот (то есть более крупных генов) размером до 23 тысяч пар нуклеотидов (т. п. н.). Причём, фаги без вставок — менее 38 т. п. н. или, напротив, со слишком большими вставками — более 52 т. п. н. не развиваются и не поражают бактерии[29].
Бактериофаги M13, фаг Т4, T7 и фаг λ используют для изучения белок-белковых, белок-пептидных и ДНК-белковых взаимодействий методом фагового дисплея.
Поскольку размножение бактериофага возможно только в живых клетках, бактериофаги могут быть использованы для определения жизнеспособности бактерий. Данное направление имеет большие перспективы, поскольку, одним из основных вопросов при разных биотехнологических процессах является определение жизнеспособности используемых культур. С помощью метода электрооптического анализа клеточных суспензий была показана возможность изучения этапов взаимодействия фаг-микробная клетка[30].
См. также
Примечания
- ↑ 1 2 3 Сергей Головин Бактериофаги: убийцы в роли спасителей // Наука и жизнь. — 2017. — № 6. — С. 26-33
- ↑ Félix d’Hérelles (1917). «Sur un microbe invisible antagoniste des bacilles dysentériques» (PDF). Comptes rendus Acad Sci Paris. 165: 373–5. Проверено 5 September 2010.
- ↑ Вирусы бактерий
- ↑ Бактериофаг (недоступная ссылка)
- ↑ Ackermann H.-W. // Res. Microbiol., 2003. — V. 154. — P. 245—251
- ↑ Hendrix R.W. // Theor. Popul. Biol., 2002. — V. 61. — P. 471—480
- ↑ Suttle C.A. (September 2005), Vuiruses in the sea. Nature 437:356-361.
- ↑ Шестаков С. В. Как происходит и чем лимитируется горизонтальный перенос генов у бактерий. Экологическая генетика 2007. — Т. 5. — № 2. — C. 12-24.
- ↑ 1 2 Tettelin H., Masignani V., Cieslewicz M. J., Donati C., Medini D., Ward N. L., Angiuoli S. V., Crabtree J., Jones A. L., Durkin A. S., Deboy R. T., Davidsen T. M., Mora M., Scarselli M., Margarit y Ros I., Peterson J. D., Hauser C. R., Sundaram J. P., Nelson W. C., Madupu R., Brinkac L. M., Dodson R. J., Rosovitz M. J., Sullivan S. A., Daugherty S. C., Haft D. H., Selengut J., Gwinn M. L., Zhou L., Zafar N., Khouri H., Radune D., Dimitrov G., Watkins K., O’Connor K. J., Smith S., Utterback T. R., White O., Rubens C. E., Grandi G., Madoff L. C., Kasper D. L., Telford J. L.,. Wessels M. R, Rappuoli R., Fraser C. M. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: implications for the microbial «pan-genome.» Proc. Natl. Acad. Sci. USA 2005. 102: 13950-13955
- ↑ 1 2 3 4 5 Guttman B., Raya R., Kutter E. Basic Phage Biology, in Bacteriophages: Biology and Applications, (Kutter E. and Sulakvelidze A., ed.), CRP Press, 2005 FL. — P. 29-66.
- ↑ Ковалёва Е. Н. Создание биопрепарата на основе выделенных и изученных бактериофагов Enterococcus faecalis: Дис. … канд. биол. наук. — Саратов, 2009. — 151 с.
- ↑ Ackermann H.-W. // Res. Microbiol., 2003. — V. 154. — P. 245—251.
- ↑ 1 2 3 4 Ожерельева Н. Г. Краткая Медицинская Энциклопедия, М.: изд-во «Советская Энциклопедия», 1989. — издание второе.
- ↑ Бактериофаги // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- ↑ Molecular Medical Microbiology / Yi-Wei Tang, Max Sussman, Dongyou Liu, Ian Poxton, Joseph Schwartzman. — 2 ed. — Academic Press, 2014. — Vol. 1. — P. 579. — 2216 p. — ISBN 9780123977632.
- ↑ Bacillus phage G, complete genome. GenBank.
- ↑ Virus Taxonomy. Classification and Nomenclature of Viruses. Seventh Report of the International Committee on Taxonomy of Viruses / Edited by M.H.V. van Regenmontel et al. — San Diego: Academic Press, 2000. — P. 43-53, 64-129.
- ↑ Mc Grath S and van Sinderen D (editors). Bacteriophage: Genetics and Molecular Biology. — 1st. — Caister Academic Press, 2007. — ISBN [1].
- ↑ Raya R.R., Hébert E.M. Isolation of phage via induction of lysogens. Bacteriophages: Methods and Protocols, Volume 1: Isolation, Characterization, and Interaction (Martha R.J. Clokie, Andrew M. Kropinski (eds.), 2009. — V. 501. — P. 23-32.
- ↑ 1 2 Микробиология: учеб. пособие / В. В. Лысак. — Минск: БГУ, 2007. — 430 с.
- ↑ Адамс М. Бактериофаги / М. Адамс. — М.: Медгиз, 1961. — 521 с.
- ↑ Гольдфарб Д. М., Бактериофагия / Д. М. Гольдфарб. — М.: Медгиз, 1961. — 299 с.
- ↑ BBC Horizon: Phage — The Virus that Cures 1997-10-09
- ↑ Parfitt T (2005). «Georgia: an unlikely stronghold for bacteriophage therapy». Lancet 365 (9478): 2166–7. DOI:10.1016/S0140-6736(05)66759-1. PMID 15986542.
- ↑ Thiel, Karl (January 2004). «Old dogma, new tricks—21st Century phage therapy». Nature Biotechnology (Nature Publishing Group) 22 (1): 31–36. DOI:10.1038/nbt0104-31. PMID 14704699. Проверено 2007-12-15.
- ↑ Aguita, Maria. Combatting Bacterial Infection, LabNews.co.uk. Архивировано 28 февраля 2009 года. Проверено 5 мая 2009.
- ↑ Pirisi A (2000). «Phage therapy—advantages over antibiotics?». Lancet 356 (9239): 1418. DOI:10.1016/S0140-6736(05)74059-9. PMID 11052592.
- ↑ Все юные жители Крымска привиты от гепатита A | РИА ФедералПресс
- ↑ Щелкунов С. Н. Генетическая инженерия / С. Н. Щелкунов. — Новосибирск: Сиб. унив. изд-во, 2004. — 496 с.
- ↑ Guliy O.I., Bunin V.D., O’Neil D., Ivnitski D., Ignatov O.V. A new electro-optical approach to rapid assay of cell viability // Biosensors and Bioelectronics. 2007. V. 23. P. 583—587.
Ссылки
wikipedia.green
к чему относятся эти вещества?
В конце XIX начале XX века сразу несколько ученых заметили, что в природе существуют особые микроскопические частички, которые способны замедлять, а порой и полностью прекращать рост и размножение бактерий. Эти частички получили название – бактериофаги.
Что такое бактериофаг?
Бактериофаг относится к вирусам. В своей структуре он также содержит ядро с генетическим материалом – цепочкой ДНК или реже РНК, которое окружено белковой защитной оболочкой. Фаги помимо ядра имеют еще и хвост, который обычно в два — четыре раза больше его. Вот только поражает этот вирус не птиц, животных или человека, а бактерии.
Действует он точно так же, как и любой другой вирус. Атаковав бактерию, фаг прикрепляется к её оболочке, постепенно растворяет её и впрыскивает внутрь свою ДНК, которая полностью перестраивает весь метаболизм клетки. В итоге бактерия начинает синтезировать новых фагов, которые после её гибели выходят наружу и атакуют другие клетки возбудителя.
В переводе с латыни название этого микроорганизма значит – пожирающий бактерии. Ученые считают, что бактериофаги являются самой древней и многочисленной группой вирусов.
Живут они везде, где есть бактерии. То есть их можно найти в воздухе, воде, почве, на продуктах питания и даже в организме рыб, птиц, животных и человека.
Виды и применение
На сегодняшний день Международный комитет по таксономии вирусов выделяет девятнадцать различных семейств. Бактериофаги относят к тому или иному семейству в зависимости от особенностей строения нити ДНК, ядра в котором она находится и хвоста.
Однако в медицине для удобства чаще применяется другая классификация, в зависимости от того для каких именно бактерий опасен тот или иной фаг. Так, на сегодняшний день существуют:
- Бактериофаг колипротейный, который используется для лечения воспалительных заболеваний ЛОР-органов вызванных бактериями Proteus vulgaris и mirabilis.
- Бактериофаг стафилококковый, который вызывает гибель стафилококков, причем, в отличие от антибиотиков, практически всех видов.
- Бактериофаг стрептококковый, влияет на стрептококков и особенно хорошо помогает при стрептококковых ангинах в тех случаях, когда антибиотики оказываются малоэффективными.
- Бактериофаг синегнойной палочки, способной провоцировать развитие тяжелых отитов, воспаление миндалин и длительные ринит, которые часто переходят в хронический гайморит или фронтит.
- Пиобактериофаг поливалентный – комплексный препарат, который врачи назначают при ЛОР-заболеваниях вызванных сразу несколькими возбудителями. А именно стрептококками и стафилококками разных видов, протеем, синегнойной палочкой, энтерококками, клебсиеллой или кишечной палочкой. Обычно применяется для лечения ангин, отита, гайморита и пневмоний.
- Бактериофаг клебсиел пневмонии, активен в том случае, когда воспаление в дыхательных путях вызвано клебсиеллами.
Бактериофаги практически не имеют противопоказаний, их применение разрешено и при беременности, у маленьких детей и у лиц, страдающих хроническими заболеваниями. Способ, дозировка и продолжительность приема в каждом случае определяются врачом.
Преимущества
Всего столетие назад антибиотики были единственным средством борьбы с различными видами инфекций. Однако за последние десятилетия из-за развивающейся устойчивости бактерий справиться с болезнью они могут не всегда. В таком случае врачи и могут назначить вам бактериофаг. Причем в отличие от антибиотиков у такого средства есть масса преимуществ:
- У бактериофагов практически нет противопоказаний и привыкание к ним не развивается.
- В отличие от антибиотиков они не действуют угнетающе на иммунную систему организма и не влияют на полезную микрофлору.
- Сочетаются с другими лекарственными средствами и методами лечения. При необходимости могут применяться совместно с антибиотиками.
- Могут помочь даже в случае вялотекущих, хронических инфекций, когда антибиотики не оказывают нужного действия.
Применяются бактериофаги как внутрь, так и местно, например, в виде промываний. Единственным противопоказанием к приему этого препарата может быть аллергическая реакция на компоненты, которые входят в состав.
Вконтакте
Google+
elaxsir.ru
Бактериофаги — это… Что такое Бактериофаги?
фаги, бактериальные вирусы, вызывающие разрушение (лизис) бактерий и других микроорганизмов. Б. размножаются в клетках, лизируют их и переходят в др., как правило, молодые, растущие клетки. Впервые перевиваемый лизис бактерий (сибиреязвенной палочки) наблюдал в 1898 русский микробиолог Н. Ф. Гамалея. В 1915 английский учёный Ф. Туорт описал это же явление у гнойного стафилококка, а в 1917 французский учёный Ф. Д’Эрелль назвал литический агент, проходящий через бактериальные фильтры, «Б».Строение и химический состав. Частицы многих Б. состоят из головки округлой, гексагональной или палочковидной формы диаметром 45—140 нм и отростка толщиной 10—40 и длиной 100—200 нм (рис.). Другие Б. не имеют отростка; одни из них округлы, другие — нитевидны, размером 8х800 нм. Содержимое головки состоит преимущественно из дезоксирибону клейновой кислоты (ДНК) (длина её нити во много раз превышает размер головки и достигает 60—70 мкм, эта нить плотно скручена в головке) или рибонуклеиновой кислоты (РНК) и небольшого количества (около 3%) белка и некоторых других веществ. Отросток имеет вид полой трубки, окруженной чехлом, содержащим сократительные белки, подобные мышечным. У ряда Б. чехол способен сокращаться, обнажая часть стержня. На конце отростка у многих Б. имеется базальная пластинка с несколькими шиловидными или другие формы выступами. От пластинки отходят тонкие длинные нити, которые способствуют прикреплению фага к бактерии (см. схему). Оболочки головки и отростка состоят из белков. Общее количество белка в частице фага 50—60% , нуклеиновых кислот — 40—50% . Каждый Б. обладает специфическими антигенными свойствами, отличными от антигенов бактерии-хозяина и других фагов. Имеются антигены, общие для ряда фагов (особенно содержащих РНК).
Распространение. Б. найдены для большинства бактерий, в том числе патогенных и сапрофитных, а также .для актиномицетов (актинофаги) и сине-зелёных водорослей. Встречаются Б. в кишечнике человека и животных, в растениях, почве, водоёмах, сточных водах, навозе и т. д. Б. почвенных микроорганизмов влияют на течение микробиологических процессов в почве (денитрификацию (См. Денитрификация), аммонификацию (См. Аммонификация), азотфиксацию (См. Азотфиксация)). Размножение. Б. прикрепляется своим отростком к бактериальной клетке и, выделяя фермент, растворяет клеточную стенку; затем содержимое его головки через канадец отростка переходит внутрь клетки, где под влиянием нуклеиновой кислоты фага останавливается синтез бактериальных белков, ДНК и РНК и начинается синтез нуклеиновой кислоты, а затем и белков фага. Часть этих белков — ферменты, другая часть образует оболочку зрелой частицы Б. Более мелкие, сферические фаги попадают в бактерии без участия отростка. Если клетка бактерии заражена одновременно частицами Б., различающимися между собой по ряду свойств, то среди потомства, кроме частиц, подобных родителям, будут и такие, у которых эти свойства встречаются в новой комбинации, т. к. при размножении Б. наблюдается Рекомбинация — обмен кусками нитей нуклеиновой кислоты, являющейся носителем наследственной информации. Частицы крупных фагов выходят из бактерии, разрушая её, а некоторых мелких и нитевидных — из живых бактерий (см. Вирусы). Одни Б. весьма специфичны и способны лизировать клетки только одного какого-либо вида микроорганизмов (монофаги), другие — клетки разных видов (полифаги). Б. делят на вирулентные, вызывающие лизис клетки с образованием новых частиц, и умеренные (симбиотические), которые адсорбируются клеткой и проникают в неё, но лизиса не вызывают, а остаются в клетке в латентной (скрытой) неинфекционной форме (Профаг). Культуры, содержащие латентный фаг, называются лизогенными. Лизогения передаётся потомству бактерии. Лизогенная культура может содержать 2—3 и более фагов; она, как правило, устойчива против находящихся в ней фагов (лишь небольшая часть клеток лизируется и освобождает зрелые фаги). Воздействуя на лизогенную культуру ультрафиолетовыми или рентгеновскими лучами, перекисью водорода и некоторыми другими веществами, можно значительно увеличить количество клеток, освобождающих фаг (т. н. индукция Б.). Лизогения широко распространена среди всех видов бактерий и актиномицетов. В ряде случаев многие свойства лизогенной культуры (токсичность, подвижность бактерий и др.) зависят от наличия в ней определённых профагов. Описано много мутаций Б., сопровождающихся изменением их литической активности, строения частиц и «колоний», устойчивости против неблагоприятных воздействий и другие свойств. Б. играют большую роль в изменчивости и эволюции микробов, причём механизмы воздействия их на клетку разные (см. Трансдукция). Б. могут резко изменять азотфиксирующую способность азотобактера, токсичность и антигенные свойства патогенных бактерий и др. Практическое значение Б. Некоторые фаги (одни или в сочетании с антибиотиками (См. Антибиотики)) применяли для профилактики (фагопрофилактики) и лечения (фаготерапии) ряда бактериальных инфекционных болезней человека (дизентерия, брюшной тиф, холера, чума, стафилококковые и анаэробная инфекции и др.) и животных. Однако антибиотики и другие химиотерапевтические средства оказались эффективнее фагов, в связи с чем применение их с лечебной целью сузилось. Б. успешно применяются при определении вида бактерий, актиномицетов. Б. могут вредить производству антибиотиков, аминокислот, молочных продуктов, бактериальных удобрений и в других отраслях микробиологического синтеза. Велико значение Б. для теоретических работ по генетике и молекулярной биологии.Лит.: Раутенштейн Я. И., Бактериофагия, М., 1955; Кривиский А. С., Проблемы бактериофагии, в сборнике: Актуальные вопросы вирусологии, М., 1960; Гольдфарб Д. М., Бактериофагия, М., 1961: Стент Г., Молекулярная биология вирусов бактерий, пер. с англ., М., 1965.
Библ.: Raettig Н., Bakteriophagie. 1917—1956, Тl 1—2, Stuttg., 1958; его же, Bakteriophagie. 1957—1965, Bd 1—2, Stuttg., 1967.
Я. И. Раутенштейн.
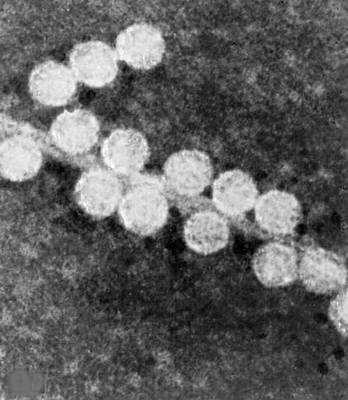
Электронная микрофотография фага МS2 без отростка, головка ок. 25 нм в диаметре.
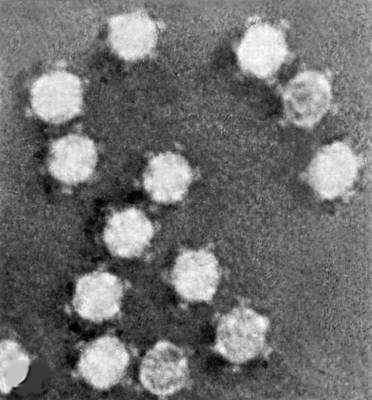
Электронная микрофотография фага Х174 без отростка с выступами, головка 25 нм в диаметре;
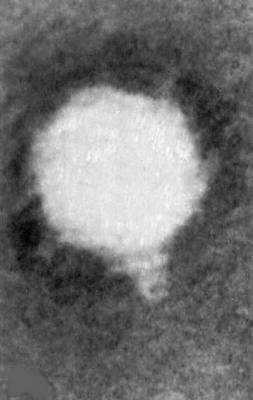
Электронная микрофотография фага Т3 с коротким отростком, головка 47х47 нм, отросток 10х15 нм
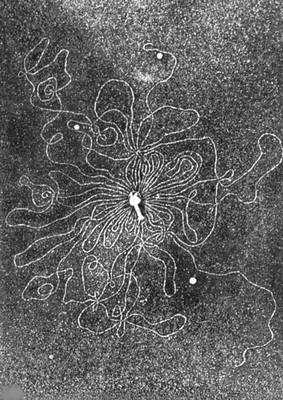
Электронная микрофотография частицы фага с освободившейся нитью дезоксирибонуклеиновой кислоты.
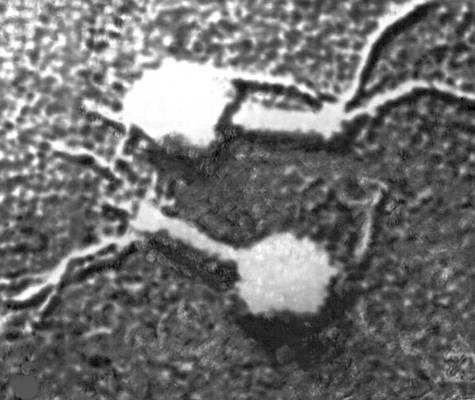
Электронная микрофотография фага Т2 при бо́льшем увеличении.
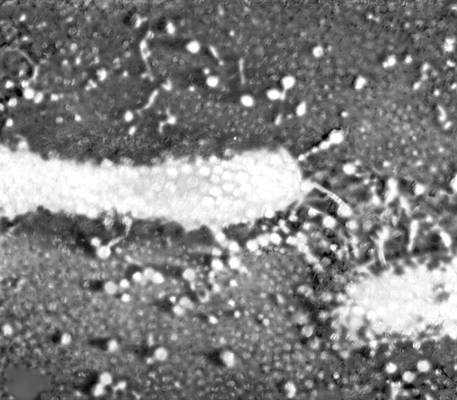
Электронная микрофотография кишечной палочки, окруженной частицами заражающего её фага Т2.
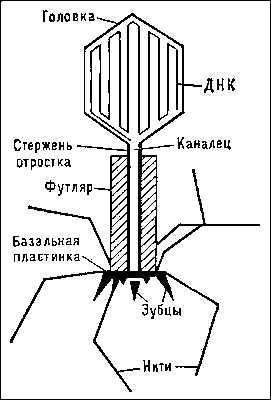
Схема строения частицы бактериофага Т2 кишечной палочки.
dic.academic.ru
Бактериофаги
Бактериофаги (вирусы бактерий)
Бактериофаги (от «бактерия» и греч. phagos — пожирающий)—вирусы бактерий, специфически проникающие в бактерии, паразитирующие в них вплоть до гибели (лизиса) бактериальной клетки.
Строение бактериофагов изучают с помощью электронной микроскопии образцов, контрастированных напылением металлов или фосфорно-вольфрамовой кислотой.
В зависимости от формы и структурной организации фаги подразделяют на несколько морфологических типов:
нитевидные;
мелкие кубические (некоторые из них имеют аналоги отростков), без отростка, с коротким отростком, с длинным отростком;
фаги сперматозоидной формы, т. е. с кубической головкой и хвостовым отростком, имеющие сокращающийся или не сокращающийся чехол отростка.
Размеры фагов колеблются от 20 до 800 нм (нитевидный тип).
Наиболее изучены крупные бактериофаги, имеющие форму сперматозоида и сокращающийся чехол отростка, например, колифаги Т2, Т4, Т6. Они состоят из головки икосаэдрического типа размером 65—100 нм и хвостового отростка длиной более 100 нм (рис. 3.14).
Хвостовой отросток имеет внутри полый цилиндрический стержень, сообщающийся с головкой, а снаружи — чехол, способный к сокращению, наподобие мышцы. Чехол присоединен к воротничку, окружающему стержень около головки. На дистальном конце отростка имеется шестиугольная базальная ,пластинка с шипами, от которых отходят нитевидные структуры — фибриллы.
Бактериофаги содержат или ДНК, или РНК. Нуклеиновые кислоты фагов могут быть двунитевыми, однонитевыми, линейными, кольцевыми. Большинство фагов содержит двунитевую ДНК, замкнутую в кольцо.
У фагов, имеющих форму сперматозоида, одна молекула двунитевой суперспирализованной ДНК находится внутри головки и защищена капсидом. Капсид состоит из белковых молекул — идентичных полипептидных субъединиц, уложенных по икосаэдрическому (кубическому) типу симметрии. В состав головки также входит полипептид, состоящий из аспарагиновой, глутаминовой кислот и лизина. У некоторых фагов внутри головки находится внутренний гистоноподобный белок, обеспечивающий суперспирализацию ДНК. Сокращающийся чехол хвостового отростка образован также белковыми субъединицами, уложенными по спиральному типу симметрии, содержащими АТФ и ионы Са2+. У некоторых фагов (например, Т2) в дистальной части отростка содержится фермент лизоцим.
Антигенные свойства. Бактериофаги содержат группоспецифические и типоспецифические антигены, обладают иммуногенными свойствами, вызывая синтез специфических антител в организме. Антитела, взаимодействуя с бактериофагами, могут нейтрализовать их литическую активность против бактерий. По типоспецифическим антигенам фаги делят на серотипы.
Резистентность. По сравнению с вирусами человека бактериофаги более устойчивы к факторам окружающей среды. Инактивируются под действием температуры 65-70 °С, УФ-облучения в высоких дозах, ионизирующей радиации, формалина и кислот. Длительно сохраняются при низкой температуре и высушивании.
Взаимодействие фагов с бактериальной клеткой. Бактериофаги инфицируют строго определенные бактерии, взаимодействуя со специфическими рецепторами клетки.
По специфичности взаимодействия различают следующие бактериофаги:
поливалентные, взаимодействующие с родственными видами бактерий;
моновалентные, взаимодействующие с бактериями определенного вида;
типовые, взаимодействующие с отдельными типами (вариантами) бактерий данного вида.
Взаимодействие фагов с бактериями может протекать, как и у других вирусов, по:
продуктивному,
абортивному
интегративному типам.
При продуктивном типе взаимодействия образуется фаговое потомство, бактерии лизируются; при абортивном типе — фаговое потомство не образуется и бактерии сохраняют свою жизнедеятельность; при интегративном типе — геном фага встраивается в хромосому бактерии и сосуществует с ней.
В зависимости от типа взаимодействия различают вирулентные и умеренные бактериофаги.
Вирулентные бактериофаги взаимодействуют с бактерией по продуктивному типу. Проникнув в бактерию, они репродуцируются с образованием 200—300 новых фаговых частиц и вызывают лизис бактерий. Взаимодействие бактериофага с бактерией напоминает взаимодействие вирусов человека с клеткой хозяина. Специфическая адсорбция фагов на бактериальной клетке происходит при наличии комплементарных рецепторов липопротеиновой или липополисахаридной природы в ее клеточной стенке. На бактериях, лишенных клеточной стенки (протопласты, сферопласты), бактериофаги не адсорбируются. Некоторые фаги в качестве рецепторов используют половые пили бактерий.
Фаги, имеющие хвостовой отросток, прикрепляются к бактериальной клетке свободным концом отростка (фибриллами, базальной пластинкой). Проникновение фаговой нуклеиновой кислоты в бактерию наиболее изучено у бактериофагов, имеющих отросток с сокращающимся чехлом. В результате активации АТФ чехол хвостового отростка сокращается, и стержень с помощью лизоцима, растворяющего прилегающий фрагмент клеточной стенки, как бы просверливает оболочку клетки. При этом ДНК фага, содержащаяся в его головке, проходит в форме нити через канал хвостового стержня и инъецируется в клетку, а капсид фага остается снаружи бактерии.
Некоторые мелкие кубические фаги, способные адсорбироваться на половых пилях, вводят свою нуклеиновую кислоту через канал этих пилей. ДНК нитевидных фагов проходит в бактерию вместе с одним из капсид-ных белков.
Инъецированная внутрь бактерии нуклеиновая кислота подавляет биосинтез компонентов клетки, заставляя ее синтезировать нуклеиновую кислоту и белки фага. Эти процессы схожи с репродукцией вирусов человека. После образования компонентов фага происходит самосборка частиц: сначала пустотелые капсиды головок заполняются нуклеиновой кислотой, затем сформированные головки соединяются с хвостовыми отростками. В результате изменения внутриклеточного осмотического давления и действия фагового лизоцима происходит разрушение оболочки, лизис бактерии и выход фагов из нее. Весь литический цикл от адсорбции бактериофага на бактерии до его выхода из нее занимает 20-40 мин.
Умеренные бактериофаги в отличие от вирулентных взаимодействуют с чувствительными бактериями либо по продуктивному, либо по интегративному типу. Продуктивный цикл умеренного фага идет в той же последовательности, что и у вирулентных фагов, и заканчивается лизисом клетки. При интегративном типе взаимодействия ДНК умеренного фага встраивается в хромосому бактерии, реплицируется синхронно с геномом размножающейся бактерии, не вызывая ее лизиса. ДНК бактериофага, встроенная в хромосому бактерии, называется профагом, а культура бактерий — лизогенной. Такое сосуществование бактерии и умеренного бактериофага называется лизогенией (от греч. lysis — разложение, genea — происхождение). Профаг, ставший частью хромосомы бактерии, при ее размножении передается по наследству потомкам.
Каким образом нуклеиновая кислота присоединяется к бактериальной хромосоме?После проникновения в бактерию ДНК умеренного фага приобретает форму кольца, а затем интегрирует по типу кроссинговера в строго определенную гомологичную область хромосомы клетки.
Итак, при лизогении образование фагового потомства не происходит. В основе «сдерживающего» механизма репродукции фагов лежит образование в бактерии специфического репрессора — низкомолекулярного белка, подавляющего транскрипцию фаговых генов. Биосинтез репрессора детерминируется генами профага. Наличием репрессора можно объяснить способность лизогенных бактерий приобретать иммунитет (невосприимчивость) к последующему заражению гомологичным или близкородственными фагами. Под иммунитетом в данном случае понимается такое состояние бактерии, при котором исключается процесс вегетативного размножения вышеуказанных фагов и лизис клетки. Однако термин «лизогения» отражает потенциальную возможность лизиса бактерии, содержащей профаг. Действительно, профаги некоторой части лизогенной культуры бактерий могут спонтанно (самопроизвольно) или направленно под действием ряда физических или химических факторов дерепрессироваться, исключаться из хромосомы, переходить в вегетативное состояние. Этот процесс заканчивается продукцией фагов и лизисом бактерий. Частота спонтанного лизиса бактерий в лизогенных культурах невелика (102, 10~6), т. е. не захватывает все клетки, обладающие иммунитетом. Частоту лизиса бактерий можно значительно увеличить, воздействуя на лизогенную культуру индуцирующими агентами (УФ-лучи, ионизирующее излучение, перекисные соединения, митомицин С и др.). Сам же феномен воздействия, приводящий к инактивации репрессора, называется индукцией профага. Явление индукции используют в генной инженерии. Однако спонтанный лизис лизогенных культур может нанести вред микробиологическому производству. Так, если микроорганизмы — продуценты биологически активных веществ оказываются лизогенными, существует опасность перехода фага в вегетативное состояние, что приведет к лизису производственного штамма этого микроба.
Геном профага может придавать бактерии новые, ранее отсутствовавшие у нее свойства. Этот феномен изменения свойств микроорганизмов под влиянием профага получил название фаговой конверсии (от лат. conver—sio— превращение). Конвертироваться могут морфологические, культуральные, биохимические, антигенные и другие свойства бактерий. Например, наличие профага в дифтерийной палочке обусловливает ее способность продуцировать дифтерийный экзотоксин.
Умеренные фаги могут быть дефектными, т. е. неспособными образовывать зрелые фаговые частицы ни в естественных условиях, ни при индукции. Геном некоторых умеренных фагов (Р1) может находиться в цитоплазме бактериальной клетки в так называемой плаз-мидной форме, не включаясь в ее хромосому. Такого рода умеренные фаги используют в качестве векторов в генной инженерии.
Практическое применение фагов. Бактериофаги используют в лабораторной диагностике инфекций при внутривидовой идентификации бактерий, т. е. определении фаговара (фаготипа). Для этого применяют метод фаготипирования, основанный на строгой специфичности действия фагов: на чашку с плотной питательной средой, засеянной «газоном» чистой культурой возбудителя, наносят капли различных диагностических типоспецифических фагов. Фаговар бактерии определяется тем типом фага, который вызвал ее лизис (образование стерильного пятна, «бляшки», или «негативной колонии», фага). Методику фаготипирования используют для выявления источника и путей распространения инфекции (эпидемиологическое маркирование). Выделение бактерий одного фаговара от разных больных указывает на общий источник их заражения.
По содержанию бактериофагов в объектах окружающей среды (например, в воде) можно судить о присутствии в них соответствующих патогенных бактерий. Подобные исследования проводят при эпидемиологическом анализе вспышек инфекционных болезней.
Фаги применяют также для лечения и профилактики ряда бактериальных инфекций. Производят брюшнотифозный, сальмонел-лезный, дизентерийный, синегнойный, стафилококковый, стрептококковый фаги и комбинированные препараты (колипротейный, пиобактериофаги и др). Бактериофаги назначают по показаниям перорально, парентерально или местно в виде жидких, таблетированных форм, свечей или аэрозолей.
Бактериофаги широко применяют в генной инженерии в качестве векторов для получения рекомбинантных ДНК.
Качественный метод определения фагов E.coli. Чашку Петри с питательным агаром засевают суточной бульонной культурой кишечной палочки газоном и подсушивают при 37 «С в течение 10—15 мин. Затем на поверхность газона наносят каплю фага и наклоняют так, чтобы капля стекла к противоположному краю. После суточной инкубации в термостате просматривают чашку, отмечая наличие зоны лизиса по месту стекания капли фага.
Количественный метод — определение титра фага по методу Грациа. Для постановки опыта предварительно: а) разливают питательный агар в чашки Петри, подсушивают в термостате; б) приготовленный полужидкий (0,7 %) питательный агар, разлитый по 3—4 мл в пробирки, растапливают в водяной бане. Делают 10-кратные разведения исследуемого фага (10~2—10~7 в зависимости от предполагаемого титра) в изотоническом растворе хлорида натрия. Затем 0,5 мл из последнего разведения фага (10~7) смешивают с таким же объемом суточной бульонной культуры чувствительных к фагу бактерий и выливают в пробирку с полужидким агаром, охлажденным до 45 «С. Смесь быстро выливают на поверхность агара в чашке Петри, где она застывает в виде тонкого слоя. Так же готовят смесь из следующего разведения фага (10~6) с бактериями и полужидким агаром и выливают на поверхность агара в другой чашке, затем — из разведения 10~5. После застывания второго слоя агара чашки инкубируют при 37 *С, затем подсчитывают число негативных колоний фага. Число этих колоний соответствует количеству фаговых частиц в засеянной смеси. Исходя из него, можно вычислить количество пятнообразующих единиц в 1 мл исходной суспензии фага. Эта величина, характеризующая концентрацию фага, называется его титром (табл. 5.3.1).
Определение спектра литического действия фага. Чашку с питательным агаром делят на квадраты по числу испытуемых бактериальных культур. На каждый квадрат петлей наносят каплю соответствующей бульонной культуры и распределяют ее по агару в пределах данного квадрата. Затем на каждый засеянный квадрат петлей или пастеровской пипеткой наносят по одной капле испытуемого фага. После суточной инкубации в термостате просматривают чашку, отмечая те квадраты, где имеется сплошной лизис бактерий или так называемые стерильные пятна набактериальном газоне. Количество различных бактериальных культур, которые лизируются испытуемым фагом, определяет широту спектра его литического действия.
Фаготипирование бактерий. Испытуемую суточную бульонную культуру бактерий засевают на поверхность питательного агара в чашке Петри, слегка подсушивают в термостате, затем делят на квадраты, на которые пастеровской пипеткой наносят по одной капле различных типоспецифических фагов. После суточной инкубации отмечают на чашке те квадраты, в которых имеется сплошной лизис бактерий. Фаготип бактериальной культуры определяется тем типом фага, который вызывает ее лизис.
Определение лизогении. Исследуемую суточную бульонную культуру центрифугируют для отделения фага от бактерий. В том случае, если бактерии спонтанно продуцируют фаг, последний будет содержаться в надосадочной жидкости. Для выявления фага надосадочную жидкость засевают на газон индикаторной (чувствительной) бактериальной культуры, на котором через
1 сут инкубации при 37 «С образуются очаги лизиса — «стерильные» пятна. При отрицательном результате опыта исследуемую бактериальную культуру предварительно подвергают УФ-облучению с целью индукции содержащегося в ней профага. Затем поступают так же, как и в предыдущем опыте.
8
studfile.net
Бактериофаги — WiKi
Бактериофа́ги, или фа́ги (от др.-греч. φᾰγω «пожираю») — вирусы, избирательно поражающие бактериальные клетки и клетки архей. Чаще всего бактериофаги размножаются внутри бактерий и вызывают их лизис. Как правило, бактериофаг состоит из белковой оболочки и генетического материала одноцепочечной или двуцепочечной нуклеиновой кислоты (ДНК или, реже, РНК). Общая численность бактериофагов в природе примерно равна общей численности бактерий (1030{\displaystyle 10^{30}} — 1032{\displaystyle 10^{32}} частиц)[1]. Бактериофаги активно участвуют в круговороте химических веществ и энергии, оказывают заметное влияние на эволюцию микробов и бактерий[1].
| Группа вирусов | |
|---|---|
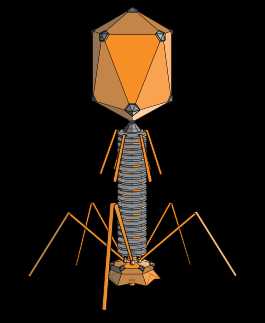 Структура типичного миовируса бактериофага. Структура типичного миовируса бактериофага. | |
| Бактериофаги | |
| не определён | |
| Домен Вирусы | |
Все вирусы, поражающие бактерий | |
История
Английский бактериолог Фредерик Туорт в статье 1915 года описал инфекционную болезнь стафилококков, возбудитель которой проходил через фильтры, и его можно было переносить от одной колонии к другой.
Независимо от Фредерика Туорта французско-канадский микробиолог Феликс Д’Эрелль 3 сентября 1917 года сообщил об открытии бактериофагов.[2] Наряду с этим известно, что российский микробиолог Николай Фёдорович Гамалея ещё в 1897 году впервые наблюдал явление лизиса бактерий (сибиреязвенной палочки) под влиянием перевиваемого агента[3][4].
После открытия явлений бактериофагии Д’Эрелль развил учение о том, что бактериофаги патогенных бактерий, являясь их паразитами, играют большую роль в патогенезе инфекций, обеспечивая выздоровление больного организма, а затем создания специфического иммунитета. Это положение привлекло к явлению бактериофагии внимание многих исследователей, которые предполагали найти в фагах важное средство борьбы с наиболее опасными инфекционными болезнями человека и животных.
Также Феликс Д’Эрелль выдвинул предположение, что бактериофаги имеют корпускулярную природу. Однако только после изобретения электронного микроскопа удалось увидеть и изучить ультраструктуру фагов. Долгое время представления о морфологии и основных особенностях фагов основывались на результатах изучения фагов группы Т — Т1, Т2,…, Т7, которые размножаются на Е. coli штамма B. Однако с каждым годом появлялись новые данные, касающиеся морфологии и структуры разнообразных фагов, что обусловило необходимость их морфологической классификации.
Роль бактериофагов в биосфере
 Бактериофаг ϕpp2 патогенных вибрионов V. parahaemolyticus
Бактериофаг ϕpp2 патогенных вибрионов V. parahaemolyticusи V. alginolyticus
Бактериофаги представляют собой наиболее многочисленную, широко распространённую в биосфере и, предположительно, наиболее эволюционно древнюю группу вирусов[5][6]. Приблизительный размер популяции фагов составляет более 1030 фаговых частиц[7].
В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, выделения человека и животных, вода и т. д.) микроорганизмами, тем в большем количестве в нём встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами чернозёмы и почвы, в которые вносились органические удобрения.
Бактериофаги выполняют важную роль в контроле численности микробных популяций, в автолизе стареющих клеток, в переносе бактериальных генов, выступая в качестве векторных «систем»[8].
Действительно, бактериофаги представляют собой один из основных подвижных генетических элементов. Посредством трансдукции они привносят в бактериальный геном новые гены. Было подсчитано, что за 1 секунду могут быть инфицированы 1024 бактерий[9]. Это означает, что постоянный перенос генетического материала распределяется между бактериями, обитающими в сходных условиях.
Высокий уровень специализации, долгосрочное существование, способность быстро репродуцироваться в соответствующем хозяине способствует их сохранению в динамичном балансе среди широкого разнообразия видов бактерий в любой природной экосистеме. Когда подходящий хозяин отсутствует, многие фаги могут сохранять способность к инфицированию на протяжении десятилетий, если не будут уничтожены экстремальными веществами либо условиями внешней среды[10].
Строение бактериофагов
1 — головка, 2 — хвост, 3 — нуклеиновая кислота, 4 — капсид, 5 — «воротничок», 6 — белковый чехол хвоста, 7 — фибрилла хвоста, 8 — шипы, 9 — базальная пластинкаБактериофаги различаются по химической структуре, типу нуклеиновой кислоты, морфологии и характеру взаимодействия с бактериями. По размеру бактериальные вирусы в сотни и тысячи раз меньше микробных клеток.
Типичная фаговая частица (вирион) состоит из головки и хвоста. Длина хвоста обычно в 2—4 раза больше диаметра головки. В головке содержится генетический материал — одноцепочечная или двуцепочечная РНК или ДНК с ферментом транскриптазой в неактивном состоянии, окружённая белковой или липопротеиновой оболочкой — капсидом, сохраняющим геном вне клетки[11].
Нуклеиновая кислота и капсид вместе составляют нуклеокапсид. Бактериофаги могут иметь икосаэдральный капсид, собранный из множества копий одного или двух специфичных белков. Обычно углы состоят из пентамеров белка, а опора каждой стороны из гексамеров того же или сходного белка. Более того, фаги по форме могут быть сферические, лимоновидные или плеоморфные[12].
Хвост, или отросток, представляет собой белковую трубку — продолжение белковой оболочки головки, в основании хвоста имеется АТФаза, которая регенерирует энергию для инъекции генетического материала. Существуют также бактериофаги с коротким отростком, не имеющие отростка и нитевидные[13].
Головка округлой, гексагональной или палочковидной формы диаметром 45—140 нм. Отросток толщиной 10—40 и длиной 100—200 нм. Одни из бактериофагов округлы, другие нитевидны, размером 8×800 нм. Длина нити нуклеиновой кислоты во много раз превышает размер головки, в которой находится в скрученном состоянии, и достигает 60—70 мкм. Отросток имеет вид полой трубки, окружённой чехлом, содержащим сократительные белки, подобные мышечным. У ряда вирусов чехол способен сокращаться, обнажая часть стержня. На конце отростка у многих бактериофагов имеется базальная пластинка, от которой отходят тонкие длинные нити, способствующие прикреплению фага к бактерии. Общее количество белка в частице фага — 50-60%, нуклеиновых кислот — 40-50%.[14]
Фаги, как и все вирусы, являются абсолютными внутриклеточными паразитами. Хотя они содержат всю информацию для запуска собственной репродукции в соответствующем хозяине, у них нет механизмов для выработки энергии и рибосом для синтеза белка. Размер известных фаговых геномов варьирует от нескольких тысяч до 498 тысяч пар оснований (геном фага G, поражающего бацилл)[15][16].
Систематика бактериофагов
Большое количество выделенных и изученных бактериофагов определяет необходимость их систематизации. Этим занимается Международный комитет по таксономии вирусов (ICTV). В настоящее время, согласно Международной классификации и номенклатуре вирусов, бактериофаги разделяют в зависимости от типа нуклеиновой кислоты и морфологии.
На данный момент выделяют девятнадцать семейств. Из них только два РНК-содержащих и только пять семейств имеют оболочку. Из семейств ДНК-содержащих вирусов только два семейства имеют одноцепочечные геномы. У девяти ДНК-содержащих семейств геном представлен кольцевой ДНК, а у других девяти — линейной. Девять семейств специфичны только для бактерий, остальные девять только для архей, а (Tectiviridae) инфицирует как бактерий, так и архей[17].
| Порядок | Семейство | Морфология | Нуклеиновая кислота | Пример |
|---|---|---|---|---|
| Caudovirales | Myoviridae | Без оболочки, сократительный хвост | Линейная дцДНК | Фаг Т4, фаг μ, PBSX, P1Puna-like, P2, I3, Bcep 1, Bcep 43, Bcep 78 |
| Siphoviridae | Без оболочки, несократительный хвост (длинный) | Линейная дцДНК | Фаг λ, фаг T5, phi, C2, L5, HK97, N15 | |
| Podoviridae | Без оболочки, несократительный хвост (короткий) | Линейная дцДНК | Фаг T7, фаг T3, P22, P37 | |
| Ligamenvirales | Lipothrixviridae | В оболочке, палочкообразные | Линейная дцДНК | Вирус »Acidianus filamentous» 1 |
| Rudiviridae | Без оболочки, палочкообразные | Линейная дцДНК | Палочкообразный вирус »Sulfolobus islandicus» 1 | |
| Неизвестен | Ampullaviridae | В оболочке, бутылкообразные | Линейная дцДНК | |
| Bicaudaviridae | Без оболочки, лемонообразные | Кольцевая дцДНК | ||
| Clavaviridae | Без оболочки, палочкообразные | Кольцевая дцДНК | ||
| Corticoviridae | Без оболочки, изометрические | Кольцевая дцДНК | ||
| Cystoviridae | В оболочке, сферические | Сегментированная дцРНК | ||
| Fuselloviridae | Без оболочки, лемонообразные | Кольцевая дцДНК | ||
| Globuloviridae | В оболочке, изометрические | Линейная дцДНК | ||
| Guttaviridae | Без оболочки, яйцевидные | Кольцевая дцДНК | ||
| Inoviridae | Без оболочки, нитевидные | Кольцевая оцДНК | M13 | |
| Leviviridae | Без оболочки, изометрические | Линейная оцРНК | MS2, Qβ | |
| Microviridae | Без оболочки, изометрические | Кольцевая оцДНК | ΦX174 | |
| Plasmaviridae | В оболочке, плеоморфные | Кольцевая дцДНК | ||
| Tectiviridae | Без оболочки, изометрические | Линейная дцДНК |
Взаимодействие бактериофага с бактериальными клетками
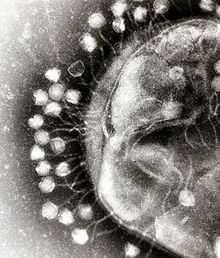 Адсорбция бактериофагов на поверхности бактериальной клетки
Адсорбция бактериофагов на поверхности бактериальной клеткиПо характеру взаимодействия бактериофага с бактериальной клеткой различают вирулентные и умеренные фаги[13]. Вирулентные фаги могут только увеличиваться в количестве посредством литического цикла[10]. Процесс взаимодействия вирулентного бактериофага с клеткой складывается из нескольких стадий: адсорбции бактериофага на клетке, проникновения в клетку, биосинтеза компонентов фага и их сборки, выхода бактериофагов из клетки[9][19].
Первоначально бактериофаги прикрепляются к фагоспецифическим рецепторам на поверхности бактериальной клетки. Хвост фага с помощью ферментов, находящихся на его конце (в основном лизоцима), локально растворяет оболочку клетки, сокращается и содержащаяся в головке ДНК инъецируется в клетку, при этом белковая оболочка бактериофага остаётся снаружи. Инъецированная ДНК вызывает полную перестройку метаболизма клетки: прекращается синтез бактериальной ДНК, РНК и белков. ДНК бактериофага начинает транскрибироваться с помощью собственного фермента транскриптазы, который после попадания в бактериальную клетку активируется. Синтезируются сначала ранние, а затем поздние иРНК, которые поступают на рибосомы клетки-хозяина, где синтезируются ранние (ДНК-полимеразы, нуклеазы) и поздние (белки капсида и хвостового отростка, ферменты лизоцим, АТФаза и транскриптаза) белки бактериофага. Репликация ДНК бактериофага происходит по полуконсервативному механизму и осуществляется с участием собственных ДНК-полимераз. После синтеза поздних белков и завершения репликации ДНК наступает заключительный процесс — созревание фаговых частиц или соединение фаговой ДНК с белком оболочки и образование зрелых инфекционных фаговых частиц[20].
Продолжительность этого процесса может составлять от нескольких минут до нескольких часов[10]. Затем происходит лизис клетки, и освобождаются новые зрелые бактериофаги[13]. Иногда фаг инициирует лизирующий цикл, что приводит к лизису клетки и освобождению новых фагов. В качестве альтернативы фаг может инициировать лизогенный цикл, при котором он вместо репликации обратимо взаимодействует с генетической системой клетки-хозяина, интегрируясь в хромосому или сохраняясь в виде плазмиды[10]. Таким образом, вирусный геном реплицируется синхронно с ДНК хозяина и делением клетки, а подобное состояние фага называется профагом. Бактерия, содержащая профаг, становится лизогенной до тех пор, пока при определённых условиях или спонтанно профаг не будет стимулирован на осуществление лизирующего цикла репликации. Переход от лизогении к лизису называется лизогенной индукцией или индукцией профага. На индукцию фага оказывает сильное воздействие состояние клетки хозяина предшествующее индукции, также как наличие питательных веществ и другие условия, имеющие место в момент индукции. Скудные условия для роста способствуют лизогенному пути, тогда как хорошие условия способствуют лизирующей реакции[10][13][20].
Очень важным свойством бактериофагов является их специфичность: бактериофаги лизируют культуры определённого вида, более того, существуют так называемые типовые бактериофаги, лизирующие варианты внутри вида, хотя встречаются поливалентные бактериофаги, которые паразитируют в бактериях разных видов[21][22].
Жизненный цикл
Умеренные и вирулентные бактериофаги на начальных этапах взаимодействия с бактериальной клеткой имеют одинаковый цикл.
- Адсорбция бактериофага на фагоспецифических рецепторах клетки.
- Инъекция фаговой нуклеиновой кислоты в клетку хозяина.
- Совместная репликация фаговой и бактериальной нуклеиновой кислоты.
- Деление клетки.
- Далее бактериофаг может развиваться по двум моделям: лизогенный либо литический путь.
- Умеренные бактериофаги после деления клетки находятся в состоянии профага (лизогенный путь).
- Вирулентные бактериофаги развиваются по литической модели:
- Нуклеиновая кислота фага направляет синтез ферментов фага, используя для этого белоксинтезирующий аппарат бактерии. Фаг тем или иным способом инактивирует ДНК и РНК хозяина, а ферменты фага совсем расщепляют её; РНК фага «подчиняет» себе клеточный аппарат синтеза белка.
- Нуклеиновая кислота фага реплицируется и направляет синтез новых белков оболочки. Образуются новые частицы фага в результате спонтанной самосборки белковой оболочки (капсид) вокруг фаговой нуклеиновой кислоты; под контролем РНК фага синтезируется лизоцим.
- Лизис клетки: клетка лопается под воздействием лизоцима; высвобождается около 200—1000 новых фагов; фаги инфицируют другие бактерии.
Применение
В медицине
Одной из областей использования бактериофагов является антибактериальная терапия, альтернативная приёму антибиотиков. Например, применяются бактериофаги: стрептококковый, стафилококковый, клебсиеллёзный, дизентерийный поливалентный, пиобактериофаг, коли, протейный и колипротейный и другие. В России зарегистрировано и применяется 13 медицинских препаратов на основе фагов[1]. В настоящее время их применяют для лечения бактериальных инфекций, которые не чувствительны к традиционному лечению антибиотиками, особенно в республике Грузия[23][24][25]. Обычно, применение бактериофагов сопровождается большим, чем антибиотики, успехом там, где присутствуют биологические мембраны, покрытые полисахаридами, через которые антибиотики обычно не проникают[26]. В настоящее время терапевтическое применение бактериофагов не получило одобрения на Западе, хотя и применяются фаги для уничтожения бактерий, вызывающих пищевые отравления, таких, как листерии[27]. В многолетнем опыте в объёме крупного города и сельской местности доказана необычайно высокая лечебная и профилактическая эффективность дизентерийного бактериофага (П. М. Лернер, 2010)[источник не указан 2316 дней]. В России терапевтические фаговые препараты делают давно, фагами лечили ещё до антибиотиков. В последние годы фаги широко использовали после наводнений в Крымске[28] и Хабаровске, чтобы предотвратить дизентерию.
В биологии
Бактериофаги применяются в генной инженерии в качестве векторов, переносящих участки ДНК, возможна также естественная передача генов между бактериями посредством некоторых фагов (трансдукция).
Фаговые векторы обычно создают на базе умеренного бактериофага λ, содержащего двухцепочечную линейную молекулу ДНК. Левое и правое плечи фага имеют все гены, необходимые для литического цикла (репликации, размножения). Средняя часть генома бактериофага λ (содержит гены, контролирующие лизогению, то есть его интеграцию в ДНК бактериальной клетки) не существенна для его размножения и составляет примерно 25 тысяч пар нуклеотидов. Данная часть может быть заменена на чужеродный фрагмент ДНК. Такие модифицированные фаги проходят литический цикл, но лизогения не происходит. Векторы на основе бактериофага λ используют для клонирования фрагментов ДНК эукариот (то есть более крупных генов) размером до 23 тысяч пар нуклеотидов (т. п. н.). Причём, фаги без вставок — менее 38 т. п. н. или, напротив, со слишком большими вставками — более 52 т. п. н. не развиваются и не поражают бактерии[29].
Бактериофаги M13, фаг Т4, T7 и фаг λ используют для изучения белок-белковых, белок-пептидных и ДНК-белковых взаимодействий методом фагового дисплея.
Поскольку размножение бактериофага возможно только в живых клетках, бактериофаги могут быть использованы для определения жизнеспособности бактерий. Данное направление имеет большие перспективы, поскольку, одним из основных вопросов при разных биотехнологических процессах является определение жизнеспособности используемых культур. С помощью метода электрооптического анализа клеточных суспензий была показана возможность изучения этапов взаимодействия фаг-микробная клетка[30].
См. также
Примечания
- ↑ 1 2 3 Сергей Головин Бактериофаги: убийцы в роли спасителей // Наука и жизнь. — 2017. — № 6. — С. 26-33
- ↑ Félix d’Hérelles. Sur un microbe invisible antagoniste des bacilles dysentériques (фр.) // Comptes rendus Acad Sci Paris. : magazine. — 1917. — Vol. 165. — P. 373—375. Архивировано 4 декабря 2010 года.
- ↑ Вирусы бактерий
- ↑ Бактериофаг (неопр.) (недоступная ссылка). Дата обращения 24 июня 2011. Архивировано 21 ноября 2013 года.
- ↑ Ackermann H.-W. // Res. Microbiol., 2003. — V. 154. — P. 245—251
- ↑ Hendrix R.W. // Theor. Popul. Biol., 2002. — V. 61. — P. 471—480
- ↑ Suttle C.A. (September 2005), Vuiruses in the sea. Nature 437:356-361.
- ↑ Шестаков С. В. Как происходит и чем лимитируется горизонтальный перенос генов у бактерий. Экологическая генетика 2007. — Т. 5. — № 2. — C. 12-24.
- ↑ 1 2 Tettelin H., Masignani V., Cieslewicz M. J., Donati C., Medini D., Ward N. L., Angiuoli S. V., Crabtree J., Jones A. L., Durkin A. S., Deboy R. T., Davidsen T. M., Mora M., Scarselli M., Margarit y Ros I., Peterson J. D., Hauser C. R., Sundaram J. P., Nelson W. C., Madupu R., Brinkac L. M., Dodson R. J., Rosovitz M. J., Sullivan S. A., Daugherty S. C., Haft D. H., Selengut J., Gwinn M. L., Zhou L., Zafar N., Khouri H., Radune D., Dimitrov G., Watkins K., O’Connor K. J., Smith S., Utterback T. R., White O., Rubens C. E., Grandi G., Madoff L. C., Kasper D. L., Telford J. L.,. Wessels M. R, Rappuoli R., Fraser C. M. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: implications for the microbial «pan-genome.» Proc. Natl. Acad. Sci. USA 2005. 102: 13950-13955 (неопр.) (недоступная ссылка). Дата обращения 24 июня 2011. Архивировано 29 декабря 2010 года.
- ↑ 1 2 3 4 5 Guttman B., Raya R., Kutter E. Basic Phage Biology, in Bacteriophages: Biology and Applications, (Kutter E. and Sulakvelidze A., ed.), CRP Press, 2005 FL. — P. 29-66.
- ↑ Ковалёва Е. Н. Создание биопрепарата на основе выделенных и изученных бактериофагов Enterococcus faecalis: Дис. … канд. биол. наук. — Саратов, 2009. — 151 с.
- ↑ Ackermann H.-W. // Res. Microbiol., 2003. — V. 154. — P. 245—251.
- ↑ 1 2 3 4 Ожерельева Н. Г. Краткая Медицинская Энциклопедия, М.: изд-во «Советская Энциклопедия», 1989. — издание второе.
- ↑ Бактериофаги // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- ↑ Molecular Medical Microbiology / Yi-Wei Tang, Max Sussman, Dongyou Liu, Ian Poxton, Joseph Schwartzman. — 2 ed. — Academic Press, 2014. — Vol. 1. — P. 579. — 2216 p. — ISBN 9780123977632.
- ↑ Bacillus phage G, complete genome (неопр.). GenBank.
- ↑ Virus Taxonomy. Classification and Nomenclature of Viruses. Seventh Report of the International Committee on Taxonomy of Viruses / Edited by M.H.V. van Regenmontel et al. — San Diego: Academic Press, 2000. — P. 43-53, 64-129.
- ↑ Mc Grath S and van Sinderen D (editors). Bacteriophage: Genetics and Molecular Biology. — 1st. — Caister Academic Press, 2007. — ISBN [1].
- ↑ Raya R.R., Hébert E.M. Isolation of phage via induction of lysogens. Bacteriophages: Methods and Protocols, Volume 1: Isolation, Characterization, and Interaction (Martha R.J. Clokie, Andrew M. Kropinski (eds.), 2009. — V. 501. — P. 23-32.
- ↑ 1 2 Микробиология: учеб. пособие / В. В. Лысак. — Минск: БГУ, 2007. — 430 с.
- ↑ Адамс М. Бактериофаги / М. Адамс. — М.: Медгиз, 1961. — 521 с.
- ↑ Гольдфарб Д. М., Бактериофагия / Д. М. Гольдфарб. — М.: Медгиз, 1961. — 299 с.
- ↑ BBC Horizon: Phage — The Virus that Cures 1997-10-09
- ↑ Parfitt T. Georgia: an unlikely stronghold for bacteriophage therapy (англ.) // The Lancet : journal. — Elsevier, 2005. — Vol. 365, no. 9478. — P. 2166—2167. — DOI:10.1016/S0140-6736(05)66759-1. — PMID 15986542.
- ↑ Thiel, Karl. Old dogma, new tricks—21st Century phage therapy (англ.) // Nature Biotechnology. — London UK: Nature Publishing Group, 2004. — January (vol. 22, no. 1). — P. 31—36. — DOI:10.1038/nbt0104-31. — PMID 14704699.
- ↑ Aguita, Maria. Combatting Bacterial Infection, LabNews.co.uk. Архивировано 28 февраля 2009 года. Дата обращения 5 мая 2009.
- ↑ Pirisi A. Phage therapy—advantages over antibiotics? (англ.) // The Lancet. — Elsevier, 2000. — Vol. 356, no. 9239. — P. 1418. — DOI:10.1016/S0140-6736(05)74059-9. — PMID 11052592.
- ↑ Все юные жители Крымска привиты от гепатита A | РИА ФедералПресс
- ↑ Щелкунов С. Н. Генетическая инженерия / С. Н. Щелкунов. — Новосибирск: Сиб. унив. изд-во, 2004. — 496 с.
- ↑ Guliy O.I., Bunin V.D., O’Neil D., Ivnitski D., Ignatov O.V. A new electro-optical approach to rapid assay of cell viability // Biosensors and Bioelectronics. 2007. V. 23. P. 583—587.
Ссылки
ru-wiki.org
