
кожный тест на туберкулез, отличия от реакции манту и подробная информация от EMC
Рассказывает педиатр, к.м.н. Юлия Климова
В дополнение к реакции Манту, которую используют для ежегодной массовой скрининговой диагностики туберкулеза, с 2008 года начали использовать Диаскинтест. Этот препарат является разработкой Российских специалистов НИИ молекулярной медицины Московской Медицинской Академии им. И.М. Сеченова. Диаскинтест, в отличие от реакции Манту, обладает почти 100% чувствительностью и специфичностью, что позволяет уменьшить вероятность развития ложноположительных реакций, имеющих место в 40-60% случаев проведения реакции Манту.
Хотя этот вид диагностики является разработкой российский ученых, научный интерес к ней высок и за рубежом. В Дании, Китае и Индонезии, Великобритании и Новой Зеландии ученые изучают свойства белка, входящего в состав Диаскинтест, ищут новые пути улучшения диагностики туберкулеза.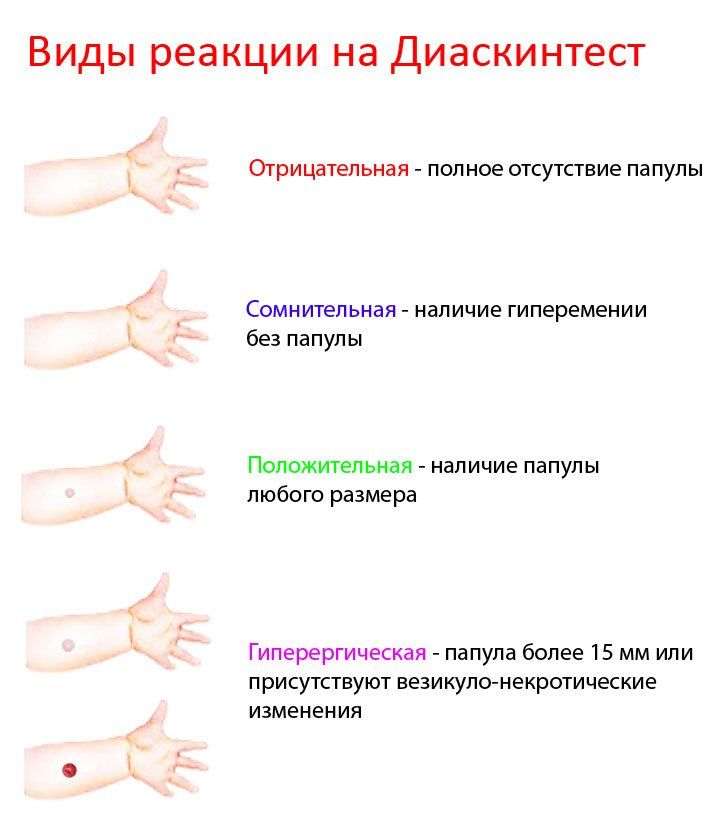
Проба подобна любым другим аллергопробам с аллергенами животного или растительного происхождения, с помощью которых диагностируют поллинозы, бронхиальную астму и другие аллергические заболевания.
О туберкулезе
В повседневной жизни нас окружает множество болезнетворных микроорганизмов. С одними мы способны легко и быстро справляться самостоятельно, против других разработана вакцинация, а третьи представляют реальную угрозу.
Несмотря на многочисленные профилактические меры, туберкулез все еще представляет угрозу здоровью людей и на территории Российской Федерации, и за ее пределами. А для людей, не вакцинированных от туберкулеза, и пациентов с первичным или приобретенным иммунодефицитом может представлять смертельную опасность.
Эта болезнь неразрывно связана с различными социальными проблемами: вооруженными конфликтами, бедностью, недоступностью медицинской помощи, с наркоманией, алкоголизмом, а также болезнью, вызываемой ВИЧ. Активная миграция населения способствует повышению риска распространения инфекции даже на благополучные по этому заболеванию районы.
Активная миграция населения способствует повышению риска распространения инфекции даже на благополучные по этому заболеванию районы.
Несмотря на снижение заболеваемости туберкулезом в России за последние годы, ситуацию все еще сложно назвать благополучной. Для начального периода болезни характерно отсутствие ярких клинических проявлений, либо они очень слабы. Это могут быть слабость, повышенная утомляемость, нарушение аппетита, раздражительность. Словом, такие состояния, которые легко недооценить и отнести к причинам, не связанным с такой грозной болезнью. Врачи называют эти проявления неспецифическими, а в случае туберкулеза — симптомами интоксикации. В этот момент особенно важно вовремя начать лечение, не допустить, чтобы болезнетворная бактерия одержала верх.
Мнение эксперта
Колпакова Людмила, врач-фтизиатр:
«Действительно, давно замечено, что бактерия туберкулеза при попадании в организм может активно размножаться и длительно не вызывать никаких симптомов.
Процесс проведения теста
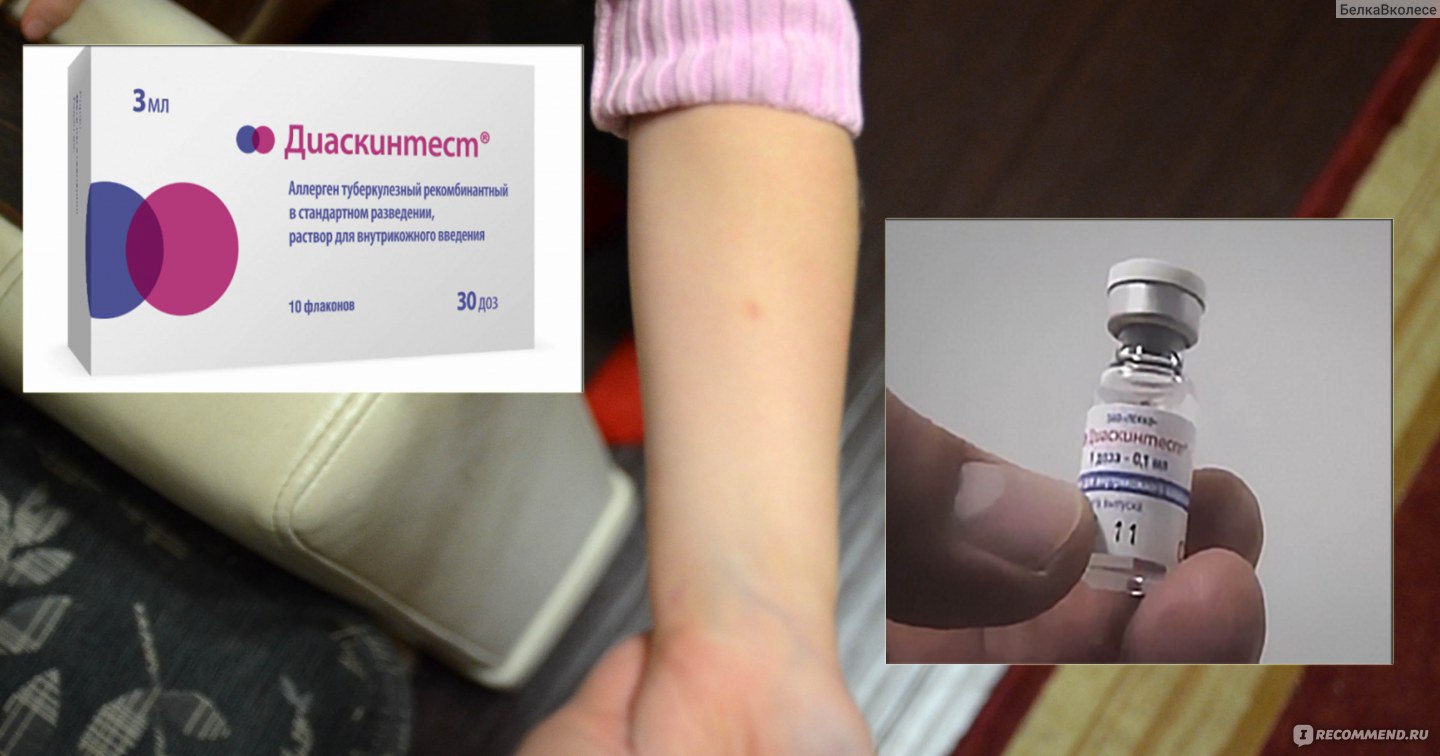
Пробу с препаратом Диаскинтест проводят аналогично тесту Манту следующим образом: внутрикожно вводят 0,1 мл препарата, в результате в месте введения образуется папула или, как ее называют иначе, «пуговка». Через 72 часа пробу оценивает педиатр или специально обученная медицинская сестра. До этого момента не следует допускать раздражения папулы нанесением различных химическх веществ, таких как гель для душа, шампунь, крем и т.п. Нельзя заклеивать или забинтовывать место пробы. На результат теста может повлиять реакция на избыточное потоотделение, к примеру, во время тренировки, или трение одежды.
До этого момента не следует допускать раздражения папулы нанесением различных химическх веществ, таких как гель для душа, шампунь, крем и т.п. Нельзя заклеивать или забинтовывать место пробы. На результат теста может повлиять реакция на избыточное потоотделение, к примеру, во время тренировки, или трение одежды.
В Детской клинике ЕМС диагностика туберкулеза с использованием Диаскинтеста проводится ежедневно, не требует специальной подготовки. Она не займет много времени, но позволит удостовериться в отсутствии опасной болезни.
Можно ли мочить манту?
Место введения как пробы манту, так и диаскинтеста, можно мочить, но нельзя тереть, чесать и применять моющие средства, такие как мыло, гель для душа и пр.
Различия реакции манту и диаскинтеста
Реакция Манту, как и Диаскинтест, является внутрикожной диагностической пробой для выявления туберкулеза. Различия состоят в составе препарата. При проведении реакции Манту используют туберкулин, который содержит частицы микобактерий человечьего и бычьего видов.
За счет чего достигается такая точность?
Диаскинтест содержит белок, включающий два антигена, характерные именно для тех видов микобактерий туберкулеза, которые могут вызывать заражение и болезнь. Важно, что этих антигенов нет в вакцинном штамме БЦЖ, который используют для вакцинации. Таким образом, результат реакции будет положительным только в том случае, если в организме имеет место размножение возбудителей туберкулеза, и будет отсутствовать ложноположительная реакция, связанная с вакцинальном штаммом микобактерий.
Показания к тесту
Согласно Приказу Минздрава России №124н от 21 марта 2017 года, Диаскинтест нужно выполнять ежегодно всем детям от 8 до 17 лет включительно. Дети из групп высокого риска, в том числе не вакцинированные против туберкулеза, нуждаются в таком обследовании дважды в год.
Противопоказания к тесту
Проведение Диаскинтеста противопоказано в период обострения любых заболеваний, в течение месяца после любой вакцинации, при эпилепсии, выраженных кожных заболеваниях, во время карантина, а также при повышенной чувствительности к любым компонентам препарата.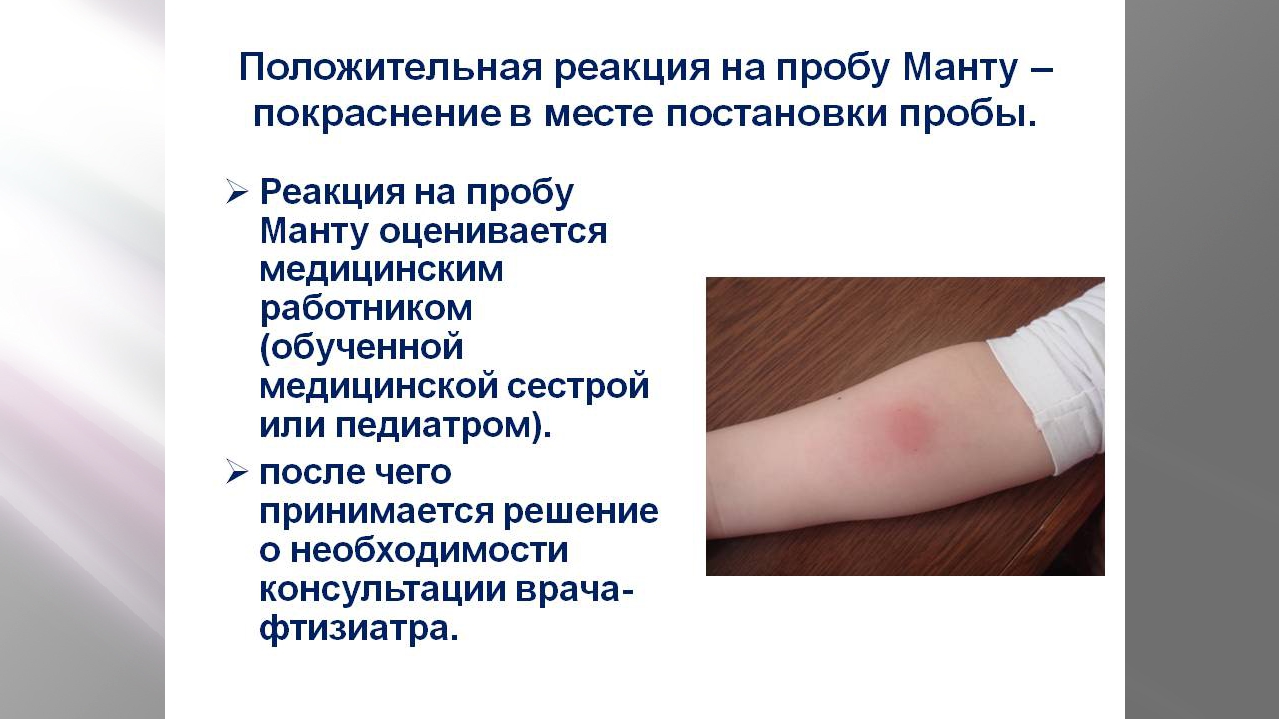
Результаты теста и их оценка
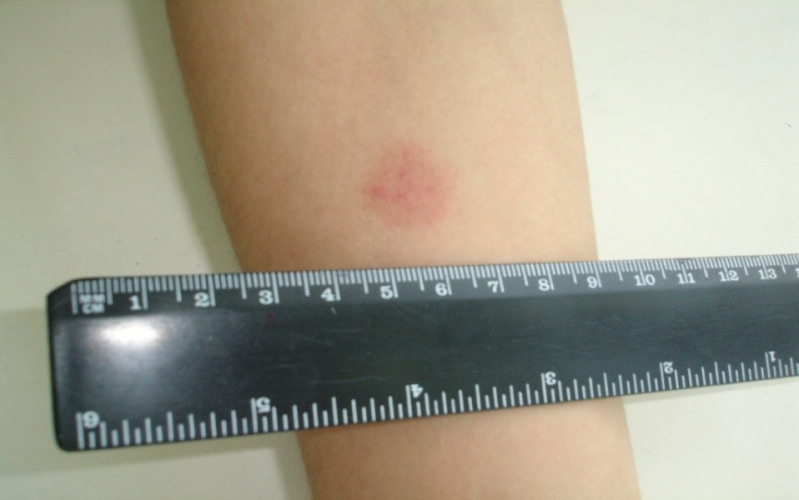
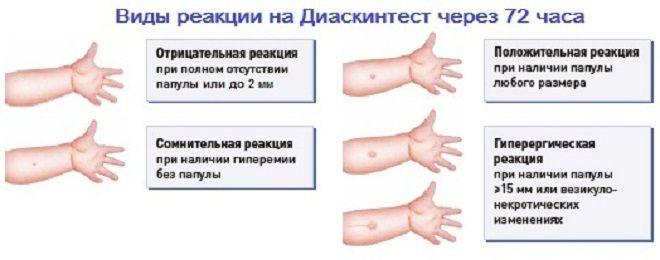
Для оценки, того, положительный или отрицательный получился тест, пробу проверяют через 72 часа после постановки путем измерения папулы прозрачной линейкой. При отрицательной реакции на коже не будет уплотнения (инфильтрата) или покраснения (гиперемии) в месте инъекции. «Уколочная» реакция до 2 мм также учитывается как отрицательная. Людям с сомнительной или положительной реакцией необходимо провести более углубленное обследование для исключения заболевания, вызванного микобактериями.
Побочные действия
Редко после введения препарата наблюдаются кратковременные реакции: повышение температуры, слабость, головная боль.
Диаскинтест – цена в Москве
Стоимость проведения диаскинтеста в Детской клинике ЕМС указана в прайс-листе по ссылке. (раздел «Врачебные манипуляции и процедуры», категория А).Консультация педиатра оплачивается отдельно.
Наши специалисты
 Так же в ЕМС взрослых и детей принимает врач-фтизиатр, который проводит дифференциальную диагностику легочных заболеваний: туберкулеза, в том числе «малых» форм и латентной туберкулезной инфекции, пневмонии, хронических неспецифических заболеваний легких, в том числе «редких» патологий: саркоидоза, альвеолитов, гистиоцитоза Х, поражений органов дыхания при иммунодефицитных состояниях.
Так же в ЕМС взрослых и детей принимает врач-фтизиатр, который проводит дифференциальную диагностику легочных заболеваний: туберкулеза, в том числе «малых» форм и латентной туберкулезной инфекции, пневмонии, хронических неспецифических заболеваний легких, в том числе «редких» патологий: саркоидоза, альвеолитов, гистиоцитоза Х, поражений органов дыхания при иммунодефицитных состояниях.
Список литературы
1. Приказ МЗ РФ №951 «Об утверждении методических рекомендаций по совершенствованию диагностики и лечения туберкулеза органов дыхания» от 29 декабря 2014 года
2. Приказ Минздрава РФ от 21.03.2017 N 124н
«Об утверждении порядка и сроков проведения профилактических медицинских осмотров граждан в целях выявления туберкулеза»
3. Слогоцкая Л.В., Сенчихина О.Ю., Никитина Г.В., Богородская Е.М. Эффективность кожного теста с аллергеном туберкулезным рекомбинантным при выявлении туберкулеза у детей и подростков Москвы в 2013 г. // Педиатрическая фармакология, 2015. – N 1. – С.99-103
// Педиатрическая фармакология, 2015. – N 1. – С.99-103
4. Bai X., Liang Y., Yang Y., Zhang J., Wu X. A new method of screening for latent tuberculosis infection: Results from army recruits in Beijing in 2014. Immunol. Lett. 2017 Jun;186:28-32. doi: 10.1016/j.imlet.2017.03.014. Epub 2017 Mar 27.
5. Aagaard C., Govaerts M., Meikle V., Vallecillo A.J., Gutierrez-Pabello J.A., Suarez-Güemes F., McNair J., Cataldi A., Espitia C., Andersen P., Pollock J.M. Optimizing antigen cocktails for detection of Mycobacterium bovis in herds with different prevalences of bovine tuberculosis: ESAT6-CFP10 mixture shows optimal sensitivity and specificity. J. Clin Microbiol. 2006 Dec;44(12):4326-35. Epub 2006 Sep 27.
6. Slogotskaya L., Bogorodskaya E., Ivanova D., Sevostyanova T. Comparative sensitivity of the test with tuberculosis recombinant allergen, containing ESAT6-CFP10 protein, and Mantoux test with 2 TU PPD-L in newly diagnosed tuberculosis children and adolescents in Moscow. PLoS One. 2018 Dec 21;13(12):e0208705. doi: .1371/journal.pone.0208705. eCollection 2018
PLoS One. 2018 Dec 21;13(12):e0208705. doi: .1371/journal.pone.0208705. eCollection 2018
В чем отличие пробы Манту и Диаскинтеста?
На территории РФ для массового обследования детей на туберкулёз используются 2 диагностические пробы:
проба Манту и Диаскинтест.
Для реакции Манту используется туберкулин, содержащий антигены, которые специфичны для палочки туберкулёза и входят в вакцинный штамм БЦЖ и БЦЖ-М. Его получают из убитой нагреванием культуры туберкулезной палочки, выращенной на крови крупного рогатого скота. Реакция Манту позволяет определить наличие иммунитета к туберкулёзу после вакцинации или при наличие этой инфекции у ребенка. При введении туберкулина развивается иммунный ответ организма в виде местной (кожной) реакции, который оценивается через 72 ч. Проводят массово детям до 7 лет включительно, старше – по показаниям.
ПРОБА МАНТУ ИСПОЛЬЗУЕТСЯ:
- для выявления раннего периода инфицирования туберкулёзом,
- для отбора детей на проведение вакцинации и ревакцинации БЦЖ (в 6-7 лет),
- для оценки состояния инфицированности туберкулёзом детского населения.
❌В случае отказа от реакции Манту в дальнейшем в 6-7 лет ребенку будет невозможно провести ревакцинацию против туберкулёза (БЦЖ), т.к она может проводиться только тем детям, у которых два года подряд реакция отрицательная. Таким образом, ребенок будет лишён дополнительной защиты от туберкулёза.
Диаскинтест получают методом генной инженерии, поэтому он не содержит следов антигенов животных. ДСТ- это комбинация 2 белков, которые характерны для патогенных штаммов палочки туберкулёза, но не входят в вакцинный штамм БЦЖ. Поэтому эта проба НЕ используется с целью отбора детей для иммунизации против туберкулёза БЦЖ. Диаскинтест, в отличие от пробы Манту, отражает чувствительность организма к белкам, которые палочка туберкулёза секретирует в активной или поздней стадии своего размножения, но не реагирует на антигены, включённые в вакцины БЦЖ и БЦЖ-М.
Проба Диаскинтест применяется для дифференциальной диагностики между поствакцинальной аллергией (после БЦЖ-М) и активным туберкулёзом, а также для диагностики туберкулёза. В соответствии с приказом 951 МЗ в нашей стране эта проба проводится с 8 лет до 17 лет включительно, а также по показаниям (по назначению фтизиатра) – детям до 7 лет включительно. В норме результат Диаскинтеста отрицательный. Дети с сомнительными и положительными результатами подлежат консультации фтизиатра для решения вопроса о дальнейшем обследовании ребёнка и его наблюдении.
В соответствии с приказом 951 МЗ в нашей стране эта проба проводится с 8 лет до 17 лет включительно, а также по показаниям (по назначению фтизиатра) – детям до 7 лет включительно. В норме результат Диаскинтеста отрицательный. Дети с сомнительными и положительными результатами подлежат консультации фтизиатра для решения вопроса о дальнейшем обследовании ребёнка и его наблюдении.
Следует отметить, что обе пробы используются для массовой диагностики туберкулёза в короткие сроки с большим охватом населения. Отклонения от нормы результатов этих проб позволяют заподозрить туберкулёз и направить пациента на дополнительное исследование (рентгенограмма, компьютерная томограмма, лабораторные исследования). Нет смысла подвергать всё детское население ежегодному рентгенологическому исследованию или другим методам исследования (Т-SPOT, квантифероновый тест, ИФА), которые назначаются по показаниям, имеют смысл у пациентов с сомнительным диагнозом, для дифференциальной диагностики туберкулеза.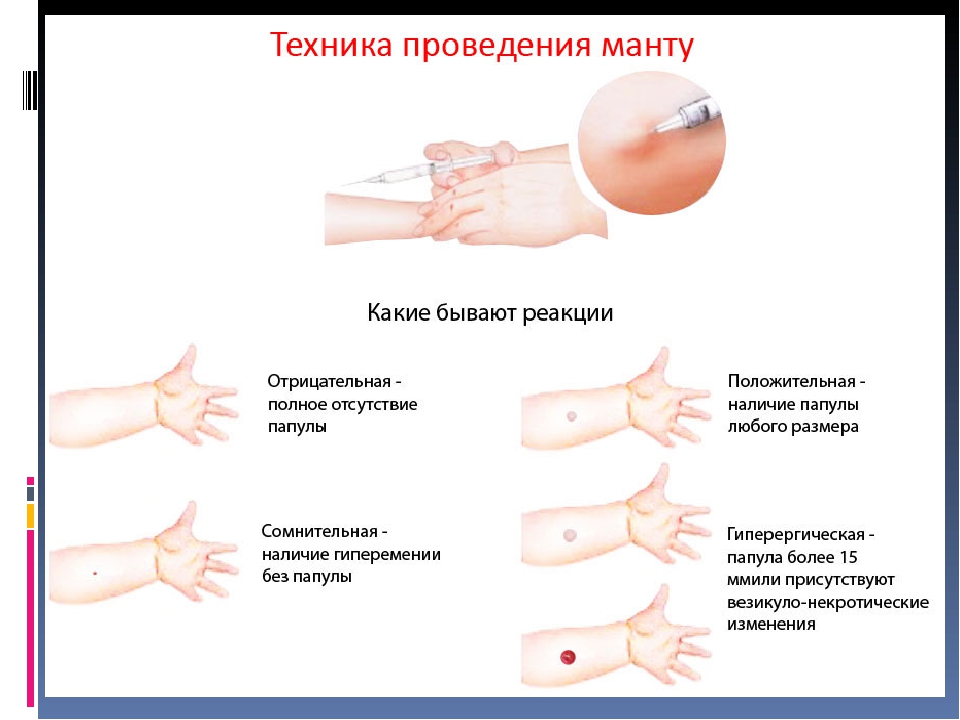 И Проба Манту, и Диаскинтест не потеряли своей актуальности в своевременной диагностике туберкулёза у детей, не смотря на появление новых современных методик.
И Проба Манту, и Диаскинтест не потеряли своей актуальности в своевременной диагностике туберкулёза у детей, не смотря на появление новых современных методик.
Цатурян Нелли Лазаревна, врач-педиатр, пульмонолог, эксперт по детским инфекциям Детской клиники «ПЛЮС»
Комментарии
Комментариев пока нет
Оставить комментарий
Диагностика туберкулеза: проба Манту или диаскинтест
В СССР диагностика туберкулёза начала проводиться туберкулином с 1965 года. Но, несмотря на такой большой опыт в туберкулинодиагностике, до сих пор продолжаются споры — какой метод лучше выбрать. Рассмотрим 2 основных метода диагностики туберкулёза у детей: проба Манту и диаскинтест.
Проба Манту
Проба Манту – это внутрикожное введение туберкулина с целью выявления туберкулёза. Несмотря на распространенное мнение – это не прививка, а тест на наличие специфических антител к микобактериям туберкулеза. Это важный метод, потому что позволяет определить туберкулёз на ранних этапах. Проведение его стандартное: 0,1 мл туберкулина вводится в среднюю треть области внутренней поверхности предплечья. Оценка результата осуществляется через 72 часа.
Это важный метод, потому что позволяет определить туберкулёз на ранних этапах. Проведение его стандартное: 0,1 мл туберкулина вводится в среднюю треть области внутренней поверхности предплечья. Оценка результата осуществляется через 72 часа.
Механизм теста следующий: он вызывает местную аллергическую реакцию замедленного типа. Сопровождается отёком, покраснением, иногда зудом. Если такая реакция возникает, значит, иммунитет к туберкулёзу сформировался, и организм уже умеет распознавать эти бактерии. Если реакции нет, то и иммунитета к микобактериям туберкулёза нет.
Существуют противопоказания для постановки пробы Манту: эпилепсия, ревматизм, опухоли, подтвержденная аллергическая реакция на туберкулин, а так же обострение аллергии, хронических инфекций, карантин в школе и детском саду. Из-за возможных аллергических реакций некоторые родители опасаются делать пробу Манту.
Диаскинтест
Существуют аналогичные методы для диагностики туберкулёза, один из них – диаскинтест.
Это синтетический препарат, что не может не радовать мам, опасающихся введения туберкулина. Так же, как и при постановке пробы Манту, возникает местная аллергическая реакция, но только в случае заболевания туберкулёзом или заражения им, которое может и не перейти в болезнь. Проведение диаскинтеста схоже с проведением пробы Манту, проверка также происходит через 72 часа.
Проводить его можно с 1 года, и одним из плюсов этого метода является отсутствие риска заразиться туберкулёзом. Этот препарат специфичен и не реагирует на другие бактерии, а также исключена реакция по поставленную ранее вакцину БЦЖ. К тому же, диаскинтестом проводят оценку эффективности лечения туберкулёза.
Детям до 8 лет, в анамнезе имеющим аллергические реакции и аллергические дерматиты, рекомендуют проводить диаскинтест. Желательно предварительно проконсультироваться у фтизиатра. В настоящее время детям с 8 лет в целях туберкулинодиагностики уже проводится диаскинтест. Но и для этого теста существуют противопоказания: обострение аллергических дерматитов, эпилепсия, обострение хронических заболеваний и острые инфекционные заболевания, за исключением подозрения на туберкулёз.
Но и для этого теста существуют противопоказания: обострение аллергических дерматитов, эпилепсия, обострение хронических заболеваний и острые инфекционные заболевания, за исключением подозрения на туберкулёз.
В любом случае, в связи со стёртой клинической картиной в начале заболевания туберкулёзом, разнообразием его форм, а также снижением социальной ответственности населения, туберкулинодиагностику необходимо проводить обязательно! А выбор метода нужно доверить грамотному специалисту, учитывая возрастные особенности, аллергоанамнез и состояние пациента на момент осмотра.
Опытные педиатры клиники «Семейный доктор» разъяснят вам различия между методами, подберут оптимальный вариант и проведут быструю и качественную диагностику в комфортных условиях. Чтобы записаться к специалисту клиники «Семейный доктор» на удобное для Вас время, позвоните по телефону единого контакт-центра в Москве +7 (495) 775 75 66, воспользуйтесь сервисом online-записи к врачу или обратитесь в регистратуру клиники.
Информацию для Вас подготовила:
Петрова Оксана Ивановна — врач-педиатр, ведущий специалист клиники, медицинский директор Детского корпуса на Усачева.
Диагностика туберкулеза у детей
Туберкулез является широко распространенным инфекционным заболеванием, имеющим высокую социальную значимость. Часто заболевание не имеет выраженных проявлений. Поэтому так важна полноценная и своевременная диагностика туберкулеза, особенно у детей.
Какие методы диагностики туберкулеза существуют сегодня?
Проба Манту
Уже более века туберкулиновая проба Манту является привычным методом выявления туберкулезной инфекции. Она используется в основном при массовой диагностике среди детей и подростков. Целью проведения туберкулиновой пробы (пробы Манту) является как раннее выявление туберкулеза, так определение необходимости повторной вакцинации здоровых детей для развития специфического иммунитета.
Суть метода заключается в том, что внутрикожно вводится специальное вещество – туберкулин – компонент микробактерий туберкулеза.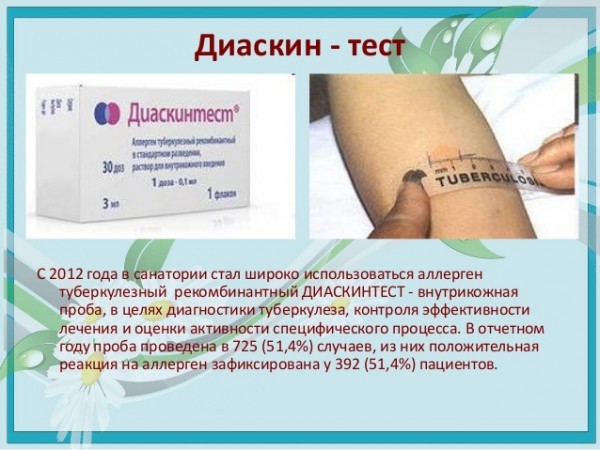 Спустя три дня медицинский работник оценивает реакцию организма на него. Проба Манту – это не прививка. Этот особый тест показывает наличие иммунитета к микобактериям туберкулеза, никакого риска заражения туберкулезом от проведенной пробы нет.
Спустя три дня медицинский работник оценивает реакцию организма на него. Проба Манту – это не прививка. Этот особый тест показывает наличие иммунитета к микобактериям туберкулеза, никакого риска заражения туберкулезом от проведенной пробы нет.
Несмотря на широкую распространенность этого метода, его дешевизну и простоту, возможность большого охвата населения, имеется ряд недостатков:
Необходим особый уход за местом инъекции;
Высокая частота ложноположительных или ложноотрицательных результатов, в том числе провоцируемых различными внешними и внутренними факторами.
Если ребенок был привит менее чем за месяц до пробы, переболел или было обострение какого-либо хронического заболевания, или ребенок чесал место пробы, его заклеивали пластырем, применяли антисептики – результаты могут быть искажены. Также результаты искажаются при нарушении техники проведения или качества препарата (хранили или перевозили неправильно). Реакция может возникать и при наличии в организме микроорганизмов, сходных с туберкулезными микобактериями, или при наличии аллергии, в том числе и на само вещество.
Диаскинтест
Более 10 лет назад был расшифрован геном микобактерий туберкулеза. Открытие антигенов, специфичных для M. Tuberculosis человека, позволило разработать препарат Диаскинтест® (ДСT), который представляет собой комплекс рекомбинантных белков продуцируемых специальной кишечной палочкой. Он так же, как и Манту, предназначен для внутрикожного введения. Все дети с положительной реакцией на пробу Манту, выявленные при массовых обследованиях, при наличии соответствующих показаний направляются на консультацию к детскому фтизиатру в противотуберкулезную организацию для диагностики активного туберкулеза и виража пробы Манту, где им в обязательном порядке делают внутрикожную пробу с ДСТ. При положительной реакции на ДСТ всех детей тщательно обследуют, в том числе с применением компьютерной томографии.
Диаскинтест:
позволяет уточнять и отсеивать ложноположительные реакции на пробу Манту,
позволяет исключить некоторые ошибки диагностики туберкулеза,
применяется в тех случаях, когда родители отказываются от пробы Манту, потому как представляет собой еще более «чистый» препарат, содержащий только белок, по своему строению идентичный белкам микобактерий туберкулеза.
Однако, он имеет ряд недостатков, так как также может давать ложноотрицательные результаты, не выявляет ранние и латентные формы туберкулеза, стоит дороже и также зависит от многих внешних и внутренних факторов, в том числе от техники введения, состояния здоровья ребенка.
Таким образом, проведение ДСТ отсеивает детей с аллергией на туберкулин, детей с ложными положительными реакциями и выявляет именно тех детей, которые подозрительны по инфицированию туберкулезом или которые уже болеют им.
Квантифероновый тест
Один из современных методов лабораторной диагностики туберкулеза, который помогает в выявлении как активного, так и скрытого туберкулеза. При его проведении никаких внутрикожных проб не ставят, а просто забирают венозную кровь на анализ, как при любом другом лабораторном исследовании. Этот метод выявляет в крови ребенка специфический интерферон, который может обнаружиться только у инфицированных туберкулезом.
Преимуществами теста являются его высокая чувствительность и специфичность, отсутствие каких-либо противопоказаний к проведению, он может проводиться вне зависимости от вакцинаций и болезней ребенка.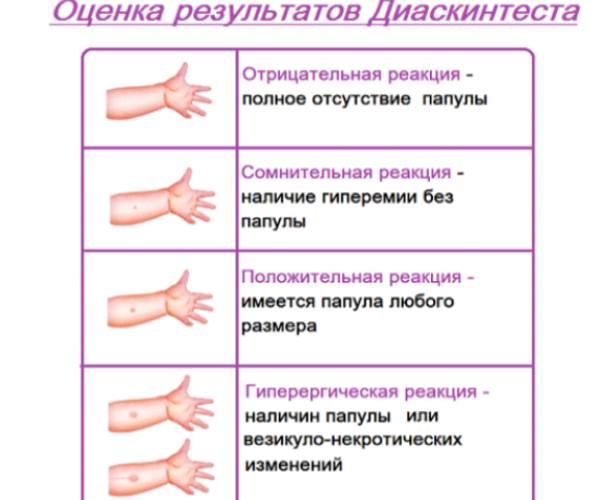 Результаты анализа не зависят от вакцинации БЦЖ.
Результаты анализа не зависят от вакцинации БЦЖ.
Квантифероновый тест делают в тех случаях, когда требуется выявление туберкулёзной инфекции после получения сомнительных результатов пробы Манту или ДСТ. Метод может использоваться для выявления туберкулёза в детских учреждениях во время карантина (когда использование внутрикожных проб недопустимо). Также тест применяют для выявления инфекции у лиц из групп риска — медицинских работников отделений ВИЧ-инфицированных, противотуберкулезных организаций, тюремных больниц, людям с противопоказаниями для туберкулиновой пробы.
Существенным недостатком теста является то, что он не может отличить обычное инфицирование и болезнь. Большинство взрослых людей в течении жизни, так или иначе, встречаются с возбудителем туберкулеза, хотя болезнь не развивается. Кроме того квантифероновый тест в 10% случаев дает ложноотрицательные результаты (при инфицировании особым штаммом «коровьего» туберкулеза). Тест довольно дорогой, делают его далеко не во всех лабораториях.
Наряду с кванифероновым тестом применяются и такие высокоточные лабораторные исследования, как ПЦР диагностика или анализ Т-клеточного маркера туберкулеза. В целом, они обладают теми же достоинствами и недостатками, что и квантифероновый тест и их используют для уточнения диагноза.
Теперь мы видим, что нет идеального единого метода диагностики. Диагностику туберкулеза проводят комплексно и поэтапно. Это позволяет выявить детей, которым необходима консультация фтизиатра, профилактическое лечение или полноценная противотуберкулезная помощь.
право выбора и право отказа / Авторские материалы / Радиостанция «Вести FM» Прямой эфир/Слушать онлайн
Манту — на выбор. Знаменитый иммунный тест на выявление заражения туберкулезом, по мнению детского омбудсмена Анны Кузнецовой, не должен быть обязательным при приеме малышей в детские сады и школы. Она не призывает совсем отказываться от обследования, но тестирование может быть иным — без укола. Хотя за него придется немало заплатить. Мнения профильных врачей о ранней диагностике опасного заболевания выяснял корреспондент «Вестей ФМ» Сергей Артемов.
Мнения профильных врачей о ранней диагностике опасного заболевания выяснял корреспондент «Вестей ФМ» Сергей Артемов.
Споры о реакции Манту — нужна она детям или нет — идут с 2013 года, когда вышли санитарные правила. Они обязали всех детей проходить тестирование на туберкулез перед поступлением в ясли и школы. И хотя копья вокруг пуговок на предплечье ломали не сами дети, но их родители — врачи поступали, и будут поступать по своим инструкциям, говорит главный фтизиопедиатр Минздрава России, доктор медицинских наук Валентина Аксенова.
«Ситуация такая в настоящее время: все дети в возрасте до 7 лет проходят ежегодное обследование пробы Манту. После 7 лет у них более информативная проба — диаскинтест. Если родители отказываются проводить либо ту, либо другую пробу — то давайте берите кровь и используйте другие методы. Не далее, как 24 марта были утверждены на профильной комиссии Российского общества фтизиатров другие тесты — альтернативные, которые можно делать, забрав кровь у ребенка».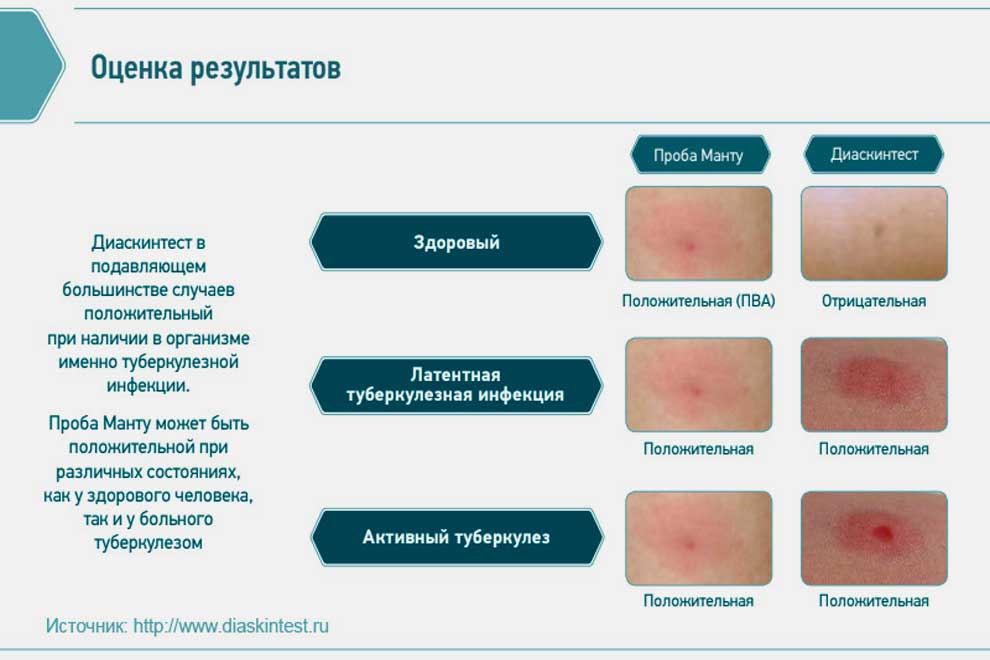
И тест Манту, и Диаскинтест проводятся уколом в руку. Лучше всего, если оба варианта будут работать в паре, полагает ведущий научный сотрудник Центрального НИИ туберкулеза доктор медицинских наук Марина Губкина.
«Все тесты любят сравнивать с позиции чувствительности и специфичности. Проба Манту — более чувствительная… Диаскинтест — более специфичный. Просто, традиционно в России, до появления Диаскинтеста, сложилась определенная тактика обследования детей на туберкулезную инфекцию — проведение пробы Манту ежегодно. Вот этой папулы, или пуговкой ее называют. Если она стала больше, или была отрицательная, а стала положительная, то отбирают в группу риска для дообследования с использованием рентгенологического метода».
Марина Губкина ссылается и на приказ Минздрава, который рекомендует как раз оба вида проверки.
«У нас уже есть новый нормативный документ, приказ №951, который определяет использование Диаскинтеста с 8 лет ежегодно, а пробы Манту с 12 месяцев до 7 лет включительно».
Возраст диагностических инъекций, как видно отличается. И зависит он от точности показаний, пояснила Валентина Аксенова.
«Диаскинтест имеет очень высокую специфичность. Если он положительный — на 100% надо идти к врачу. Значит в организме развивается уже вирулентная микобактерия. Это очень опасный вариант. Он требует обследования на туберкулез. Таких детей в школу допускать нельзя. Проба Манту малоспецифичная. У нее специфичность — около 40%. Она может давать ложноположительные результаты на аллергию, глистную инвазию, и много таких моментов. Особенно, если ребенок аллергик».
То есть, на практике распухшая пуговка Манту может показать и наличие туберкулезной инфекции, и очень мощный иммунитет ребенка, который тут же активно подавляет даже ослабленные микобактерии, и то, что у ребенка могут быть глисты или аллергия. А поскольку Диаскинтест выявляет только туберкулез, то маленьким детям, после прививок — его проводить бессмысленно, реакция покажет, что ребенок — якобы болен именно туберкулезом. Многие родители настроены против уколов их чадам. Они по-разному аргументируют свою позицию, говорит Валентина Аксенова.
Многие родители настроены против уколов их чадам. Они по-разному аргументируют свою позицию, говорит Валентина Аксенова.
«Они думают, что можно заразить. Раньше, когда были шприцы многоразовые — они боялись ВИЧ-инфекции, но сейчас это в прошлом. Шприцы теперь одноразовые. Плюс — они боятся, что у ребенка на введенное вещество будет аллергия. Мы таких детей знаем — у кого бывают аллергические реакции. Прежде, чем он сделает реакцию Манту — у него давным-давно будет написано в медицинской книжке, что ему противопоказана проба Манту».
В последние годы во многих странах мира распространяется другой принцип диагностики — без уколов, но с анализом крови, рассказывает Марина Губкина.
«Есть квантифероновый тест. Ребенку не вводят препараты, а забирают кровь и потом в лаборатории проводят исследования».
Квантифероновый тест использовался и в России — но лицензия на препарат истекла и теперь единственной альтернативой инъекционным тестам осталась так называемая Ти-спот-диагностика. Родители могут настаивать на ней — если они категорически против Манту. Врачи обязаны пойти им навстречу. Уже изданы клинические рекомендации такого тестирования. Одна проблема, замечает Валентина Аксенова — делается этот тест за собственные деньги и далеко не в каждом населенном пункте:
Родители могут настаивать на ней — если они категорически против Манту. Врачи обязаны пойти им навстречу. Уже изданы клинические рекомендации такого тестирования. Одна проблема, замечает Валентина Аксенова — делается этот тест за собственные деньги и далеко не в каждом населенном пункте:
«Такой тест делается в крупных центрах. Там, где есть иммунологические лаборатории. Намного проще сделать Диаскинтест. Те же, кто против прививок и манипуляций с ребенком, пусть едут в областной центр и делают альтернативный метод».
Тесты на туберкулез через анализ крови уже стали самыми популярными в США. А вот в Германии, например, детей при поступлении в школу вообще не проверяют на эту болезнь. В России от диагностики Минздрав отказываться не намерен. Но будут внесены поправки в вакцинацию БЦЖ — первая прививка младенцам сохранится, а вот вторую в 7-летнем возрасте, скорее всего, отменят. Тогда самый точный диаскинтест можно будет использовать уже в 5 лет.
Медики подчеркивают — они не отнимают у родителей право беспокоиться за своего ребенка, ставить ему прививки, делать диагностику — или нет.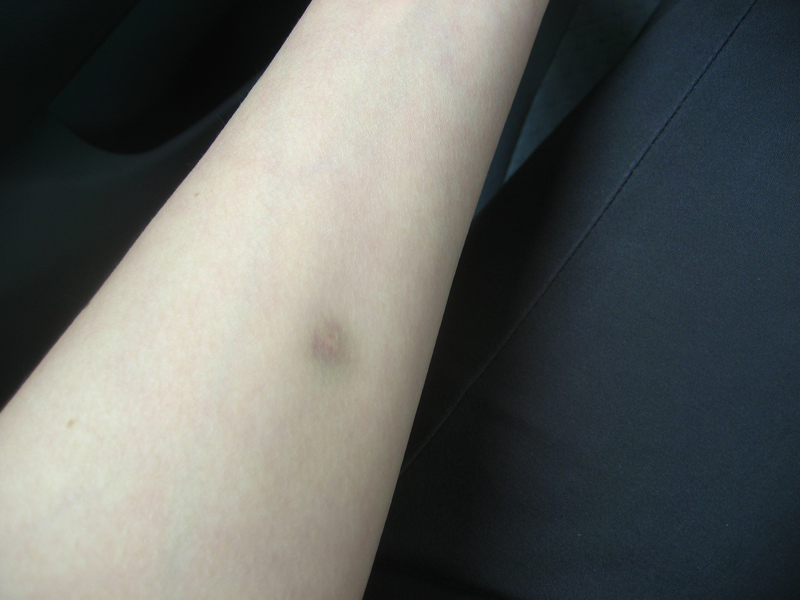 Но до тех пор, пока на врачах лежит государственная ответственность за профилактику туберкулеза — в коллективы, будь то в детские сады, или в школы — потенциальных носителей инфекции они, как и прежде, не допустят.
Но до тех пор, пока на врачах лежит государственная ответственность за профилактику туберкулеза — в коллективы, будь то в детские сады, или в школы — потенциальных носителей инфекции они, как и прежде, не допустят.
Диаскинтест / Вакцинопрофилактика / Услуги
Диаскинтест – современный препарат, позволяющий выявить латентные формы туберкулеза. В сравнении с пробой Манту этот способ первичной диагностики позволяет практически без ошибок выявлять инфицированность пациента туберкулезными бактериями. Тест может определить присутствие палочек Коха в организме еще на стадиях, когда опасная болезнь себя пока не проявила.
ЧТО ТАКОЕ ДИАСКИНТЕСТ
Как и проба Манту, Диаскин не является прививкой. Это тестовая проба, которая показывает реакцию на туберкулез в активной, неактивной форме. Для проведения иммунологического теста под кожу вводят белковые аллергены или антигены. С их помощью устанавливается иммунный ответ организма. Если он положительный, пациент находится на активной стадии болезни или инфицирован.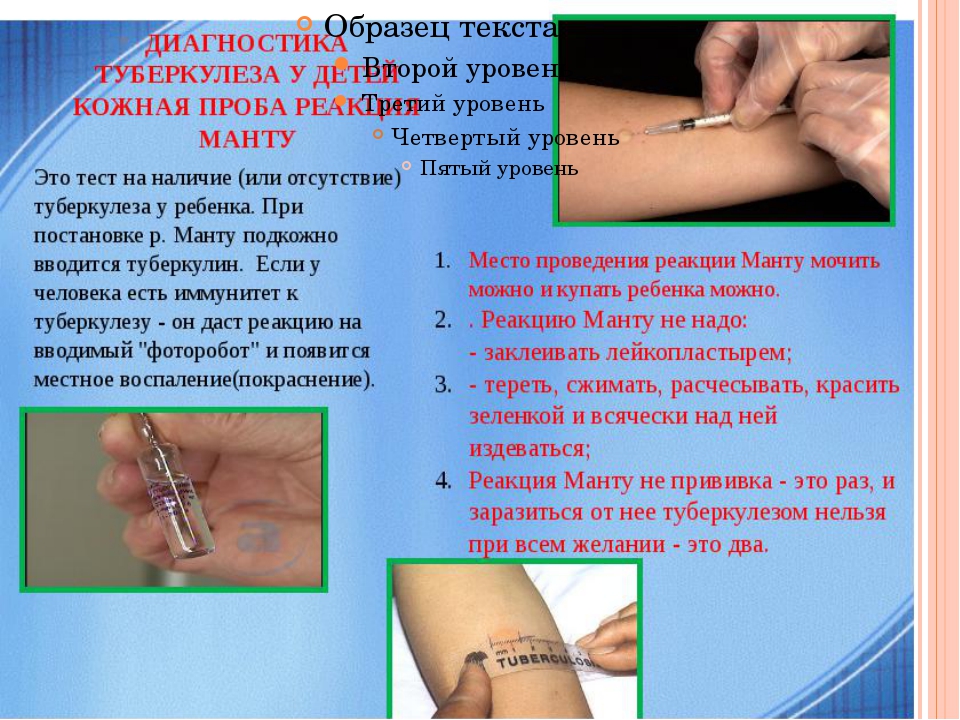 Положительная реакция – подтверждение того, что иммунитет человека знаком с антигенами.
Положительная реакция – подтверждение того, что иммунитет человека знаком с антигенами.
КАК ПРОВОДЯТ ТЕСТ
Диаскинтест выполняют таким же способом, как и остальные виды проб. Препарат вводят в область предплечья на любую руку. Обычно для правшей пробу ставят в левую руку, чтобы минимизировать механическое раздражение. Важно, чтобы человек не расчесывал зону введения препарата. Проба вводится внутрикожно тонкой иглой.
ДЛЯ ЧЕГО НУЖЕН ДИАСКИН
Многие люди путают Диаскинтест с вакциной. Но это совсем не так. Отказываться от проведения пробы не стоит, поскольку до 95 % взрослых людей сегодня инфицированы палочкой Коха. То есть, они являются носителями туберкулезных бактерий. Это носительство называют латентной формой заболевания – когда человек не болен, но болезнь может развиться при определенных условиях. Такой вариант наблюдается у 1 % пациентов. Причем инфекционный процесс долго может протекать бессимптомно. Без проведения проб выявить его только по внешним признакам невозможно.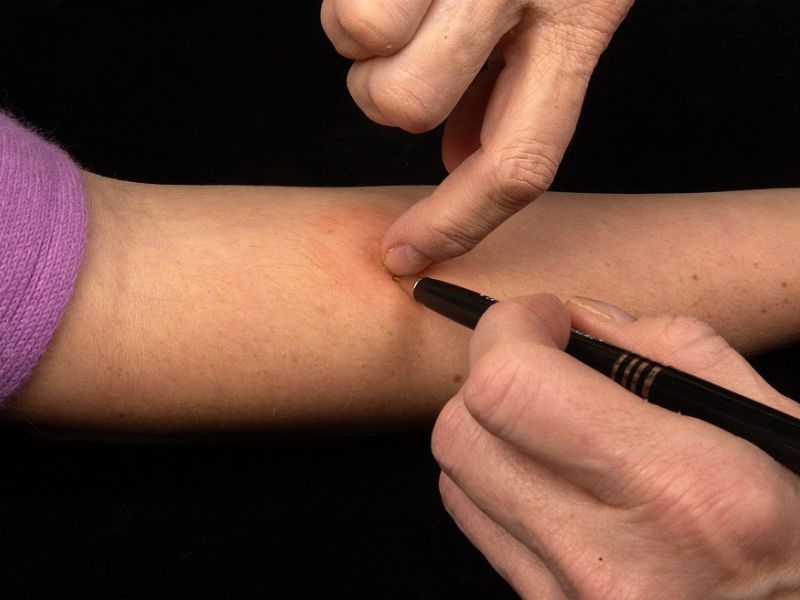 И чем раньше будет выявлен туберкулезный процесс, тем выше у пациента будут шансы на выздоровление.
И чем раньше будет выявлен туберкулезный процесс, тем выше у пациента будут шансы на выздоровление.
ПЕРИОДИЧНОСТЬ ВЫПОЛНЕНИЯ ПРОБЫ
Согласно рекомендации российского Министерства здравоохранения, Диаскинтест в обязательном порядке выполняют один раз в год в возрасте 8-17 лет. Решая, как часто можно ставить пробу, учитывают некоторые нюансы:
- Тест можно делать не раньше, чем через месяц после любой вакцинации.
- После выздоровления от любой инфекционной болезни должен пройти месяц.
- Если предыдущий результат пробы был отрицательным, следующий тест можно сделать через 2 месяца.
Контрольная проба для людей, стоящих на учете в противотуберкулезном диспансере, проводится раз в 3-6 месяцев.
Если у ребенка была положительной проба Манту, выполнять Диаскинтест ему можно с 1 года.
ПОДГОТОВКА К ПРОБЕ
Как и при Манту, постановка Диаскинтеста не требует подготовки. Необходимо лишь исключить наличие инфекционных заболеваний – за месяц до тестирования и на момент его проведения.
СОСТАВ ПРЕПАРАТА
Препарат содержит:
- Белки бактерий туберкулеза CFP10 ESAT6, подготовленные специальным образом.
- Фенол как консервант.
- Полисорбат как стабилизатор.
- Фосфорно-кислые натрий, калий.
- Хлорид натрия.
- Вода.
Диаскинтест является российским препаратом. Пройти тестирование можно в различных учреждениях. Чаще всего пробу выполняют в детских садах, школах, поликлиниках или же в противотуберкулезных институтах, центрах, диспансерах. Для взрослых обследование назначается при наличии контакта с больными или в случаях, когда человек стоит на учете в диспансере.
КАК ОЦЕНИВАЮТ РЕЗУЛЬТАТЫ
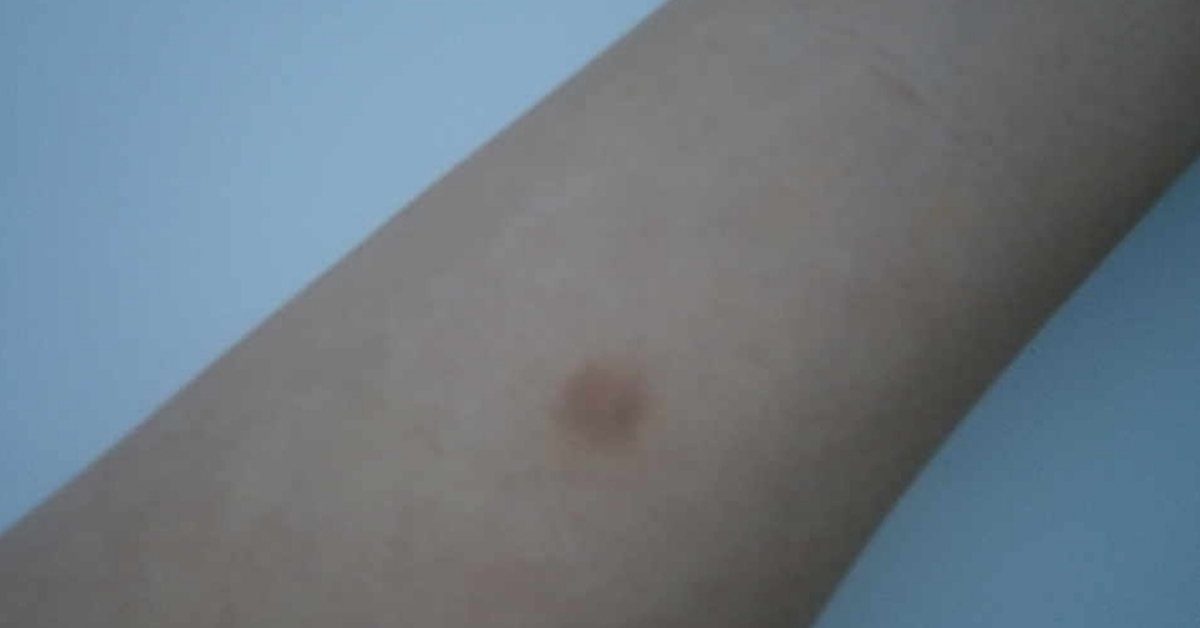
Проверка реакции выполняется на 3-4 день после проведения теста. Результат может быть положительным, отрицательным или ложноположительным (сомнительным). Отрицательный результат – норма для детей и взрослых. При полном отсутствии реакции не должно быть папул, покраснений.
Первая реакция может наблюдаться через 6 часов после введения препарата. Результат теста – ответ иммунитета человека. Воспаления, покраснения могут наблюдаться в первый же день после тестирования. Реакция может возрастать в течение 72 часов, максимальный результат наблюдается на третий день у взрослых и детей. Затем реакция угасает.
Результат теста – ответ иммунитета человека. Воспаления, покраснения могут наблюдаться в первый же день после тестирования. Реакция может возрастать в течение 72 часов, максимальный результат наблюдается на третий день у взрослых и детей. Затем реакция угасает.
ОТРИЦАТЕЛЬНЫЙ РЕЗУЛЬТАТ
Нормой при отрицательной реакции считают:
- Следы от иглы.
- Небольшой синяк на месте укола.
- Бесцветное уплотнение на месте укола размером менее 1 см.
СОМНИТЕЛЬНАЯ РЕАКЦИЯ
Ложноположительный результат устанавливается в случаях, когда у человека не появилась папула, но есть покраснения. В этих ситуациях пациенту назначают повторный тест через 2 месяца. Также необходимо пройти консультацию в противотуберкулезном диспансере, сдать флюорографию, дополнительные анализы.
ПОЛОЖИТЕЛЬНЫЙ РЕЗУЛЬТАТ
Наличие папулы любого размера говорит об инфицировании пациента туберкулезными бактериями. У взрослых размер воспалительной папулы не имеет решающего значения.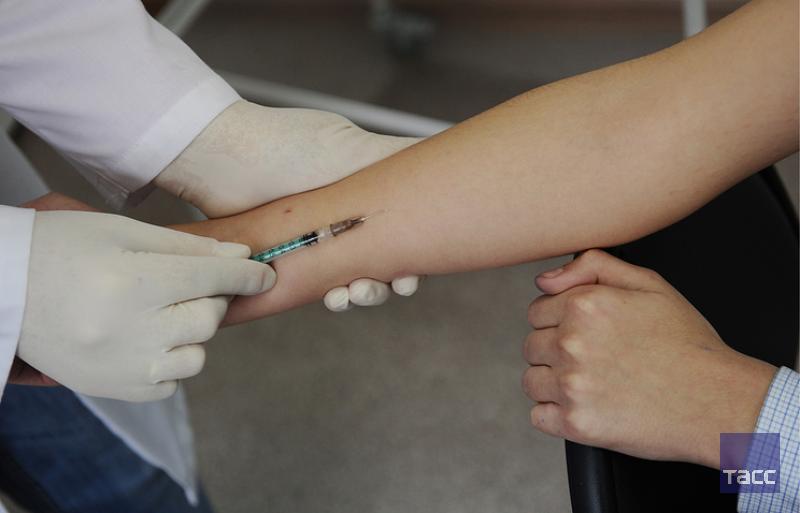 Однако для простоты оценки используется общая классификация размеров припухлостей:
Однако для простоты оценки используется общая классификация размеров припухлостей:
- Менее 5 мм – слабая.
- Не более 9 мм – умеренная.
- Свыше 10 мм – сильная.
Чем более выражена реакция, тем больше возбудителя присутствует в организме человека. Пациенту обязательно нужно обратиться в диспансер для лечения, постановки на учет.
У детей наличие папулы также требует незамедлительного обращения к врачу. Понадобятся дополнительные исследования для оценки формы заболевания – активная или латентная. Используется и специальное лечение.
ПОБОЧНЫЕ ЭФФЕКТЫ, ОСЛОЖНЕНИЯ
Диаскинтест безвреден для пациентов любого возраста, поэтому можно не опасаться реакции у детей и взрослых. Какие-либо побочные эффекты могут наблюдаться лишь в редких случаях в виде симптомов, указывающих на общую интоксикацию. К ним относят головную боль и слабость, общее недомогание, возрастание температуры. Это нормальная реакция. Не относят к поводам для беспокойства и небольшие синяки на месте укола.
Может возникать и гиперергическая реакция организма на введение препарата – образование крупной папулы диаметром более 15 мм. К такой же реакции относят сильное воспаление, раздражение ткани вокруг папулы.
ПРОТИВОПОКАЗАНИЯ К ТЕСТУ
Диаскинтест нельзя выполнять в нескольких случаях:
- Аллергии.
- Вакцинация любыми прививками в течение месяца до пробы.
- Любые заболевания в фазе обострения, острые заболевания.
- Эпилепсия.
- Заболевания кожи.
Диаскинтест, как и Манту, нельзя ставить при простуде, ОРВИ. Аллергические реакции на препарат являются противопоказанием к тестированию.
ПОВЕДЕНИЕ ПОСЛЕ ПРОБЫ
После постановки Диаскинтеста можно мочить руку, принимать душ или ванну. Категорически не рекомендуется расчесывать место укола, заклеивать эту зону пластырем, обматывать бинтом. Также не стоит смазывать место укола кремами, моющими и другими косметическими средствами. Ограничений по питанию не существует.
Диаскинтест имеет высокую чувствительность – около 80 %, а точность достигает почти 100 %. Но, как и любая медицинская тестовая методика, и эта проба может ошибаться.
Но, как и любая медицинская тестовая методика, и эта проба может ошибаться.
Диаскинтест в разы точнее Манту, поскольку он не показывает положительные реакции у людей с приобретенным противотуберкулезным иммунитетом. Вот почему эта проба считается наиболее достоверной и безопасной. С ее помощью можно узнать, не болен ли пациент на данный момент, проанализировать, не заболеет ли человек через 1-2 года.
Цены
Уважаемые посетители нашего сайта, так как прайс регулярно обновляется (вынуждены заметить, что довольно часто в сторону уменьшения), цены на все услуги размещены на отдельной странице, воспользовавшись поиском по которой, вы легко сможете найти цены на интересующие услуги. Мы стараемся пристально следить за актуальностью цен на сайте!
Также вы можете просто позвонить нашим специалистам и они быстро вас проконсультируют:
+7 (495) 577-09-41
Проверка на туберкулёз: какой метод выбрать?
Туберкулёз в России распространён настолько, что к сорока годам 70-90% жителей нашей страны инфицированы им. Это не значит, что они больны: после инфицирования иммунитет большинства людей справляется с бактерией. Вероятность заболеть составляет в среднем 8% в первые два года после инфицирования, затем постепенно снижается, и формируется приобретенный клеточный иммунитет. Среди взрослых чаще болеют люди ослабленные, живущие в плохих условиях, при недостатке света и свежего воздуха. Тем не менее, заразиться может абсолютно каждый из нас. Поэтому так важно проходить регулярные проверки на туберкулёз и проверять своих детей.
Это не значит, что они больны: после инфицирования иммунитет большинства людей справляется с бактерией. Вероятность заболеть составляет в среднем 8% в первые два года после инфицирования, затем постепенно снижается, и формируется приобретенный клеточный иммунитет. Среди взрослых чаще болеют люди ослабленные, живущие в плохих условиях, при недостатке света и свежего воздуха. Тем не менее, заразиться может абсолютно каждый из нас. Поэтому так важно проходить регулярные проверки на туберкулёз и проверять своих детей.
Зачем нужна прививка БЦЖ?
ВОЗ рекомендует массовую вакцинацию новорожденных против туберкулёза во всех странах, где эта болезнь распространена. Поэтому на 3-5 день после рождения, еще в роддоме, всем новорожденным бесплатно делают прививку БЦЖ. Но эта вакцина не защищает от инфицирования туберкулёзом. Её задача несколько иная. Если не привитый ребенок младше 2 лет будет инфицирован туберкулёзом, у него крайне высока вероятность развития туберкулезного менингита и генерализованных форм туберкулеза, которые очень быстро приводят к смерти. БЦЖ достаточно надёжно защищает ребенка именно от этих форм. Она также защищает детей от легочной формы туберкулеза, но уже менее эффективно.
БЦЖ достаточно надёжно защищает ребенка именно от этих форм. Она также защищает детей от легочной формы туберкулеза, но уже менее эффективно.
Какие ещё прививки нужны детям до 2-х лет?
Проба Манту
Проба Манту ежегодно проводится всем детям до 7 лет. Это не прививка (!), а иммунологический тест, показывающий, есть ли в организме возбудитель туберкулеза. При этом внутрикожно вводится туберкулин — специфический белок, содержащий антигены человеческого и бычьего туберкулеза. Через 72 часа измеряется размер папулы. На результат реакции Манту может повлиять посещение сауны, долгое принятие ванны, бассейн и расчёсывание места введения пробы. Временный контакт с водой никак не влияет на результаты пробы Манту, поэтому мнение о том, что Манту нельзя мочить — миф.
Диаскинтест тест
Он проводится детям с 8 лет. Также является кожной пробой. Но если в Манту используются антигены как человеческого, так и бычьего туберкулеза, то в диаскинтесте — только антигены туберкулеза человека.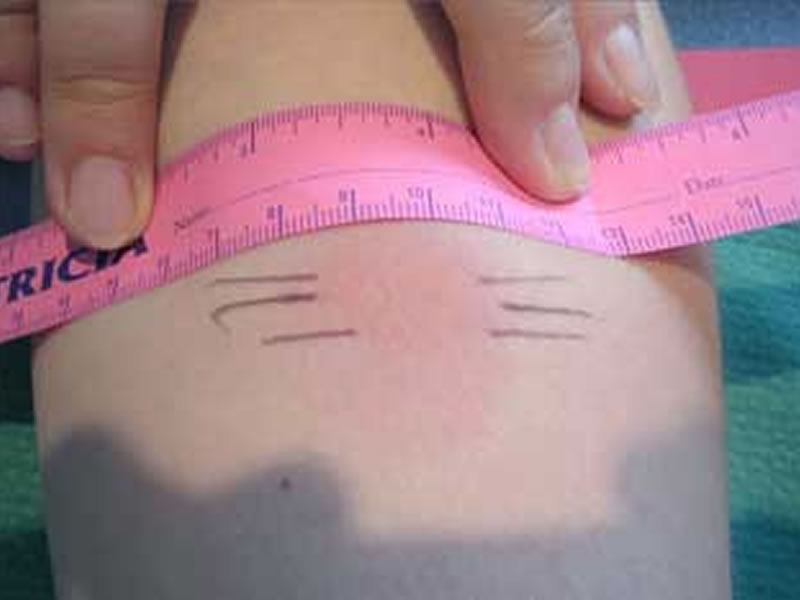 Диаскинтест гораздо более специфичен: реакция на него возникает, только если в организме есть активные бактерии туберкулеза. Проще говоря — если проба Манту положительная, это ещё ничего не значит: эта проба часто дает и ложноположительные реакции. А вот после диаскинтеста появилась реакция — это более серьёзный показатель.
Диаскинтест гораздо более специфичен: реакция на него возникает, только если в организме есть активные бактерии туберкулеза. Проще говоря — если проба Манту положительная, это ещё ничего не значит: эта проба часто дает и ложноположительные реакции. А вот после диаскинтеста появилась реакция — это более серьёзный показатель.
Точность Манту составляет 50-70%, диаскинтеста — 90%.
Квантифероновый тест
Квантифероновый тест — один из новых методов диагностики туберкулёза по анализу крови. Он основан на определении в крови гамма-интерферона, который вырабатывают клетки в ответ на внедрение туберкулезной палочки. В отличие от Манту и диаскинтеста, квантифероновый тест не требует введения в организм никаких антигенов и бактерий. Кровь для исследования берут из вены. Результат готов через несколько дней, и его точность намного выше, чем у кожных проб. Тест будет положительным, только если человек действительно болен.
T.SPOT.TB («ти-спот»)
Этот метод диагностики туберкулеза по крови аналогичен квантифероновому тесту. Отличие в том, что квантифероновый тест определяет в крови гамма-интерферон, вырабатываемый в ответ на внедрение туберкулезной палочки, а Т-СПОТ определяет сами Т-клетки, которые вырабатывают гамма-интерферон на присутствие микобактерии туберкулеза. Оба теста одинаково чувствительны и информативны (до 97%), они чувствительнее диаскинтеста и намного чувствительнее реакции Манту. «Ти-спот» практически исключает ложноположительные и ложноотрицательные реакции, в то время как квантифероновый тест все-таки может давать ложноположительную реакцию, если во время сдачи крови человек был, например, болен ОРВИ.
Отличие в том, что квантифероновый тест определяет в крови гамма-интерферон, вырабатываемый в ответ на внедрение туберкулезной палочки, а Т-СПОТ определяет сами Т-клетки, которые вырабатывают гамма-интерферон на присутствие микобактерии туберкулеза. Оба теста одинаково чувствительны и информативны (до 97%), они чувствительнее диаскинтеста и намного чувствительнее реакции Манту. «Ти-спот» практически исключает ложноположительные и ложноотрицательные реакции, в то время как квантифероновый тест все-таки может давать ложноположительную реакцию, если во время сдачи крови человек был, например, болен ОРВИ.
Сравнительная характеристика четырех тестов на туберкулёз
| Манту | Диаскинтест | Квантифероновый тест | T.SPOT.TB | |
| Точность | 50,00% | 90,00% | 97,00% | 97,00% |
| Оценка данных | Субъективная | Субъективная | Объективная | Объективная |
| Ложноположительные результаты | Часто | Редко | Крайне редко | Нет |
| При скрытой форме туберкулеза | Не достоверен | Не достоверен | Достоверен | Достоверен |
| Противопоказания | Много | Есть | Нет | Нет |
| Побочные реакции | Есть | Редко | Нет | Нет |
| При ВИЧ и других иммунодефицитах | Не информативен | Не информативен | Информативен | Информативен |
| Способ исследования | Кожный тест | Кожный тест | Анализ крови | Анализ крови |
ПЦР
Чтобы отличить активную форму туберкулеза от латентной, используется специфический тест — ПЦР (исследования мокроты на ДНК).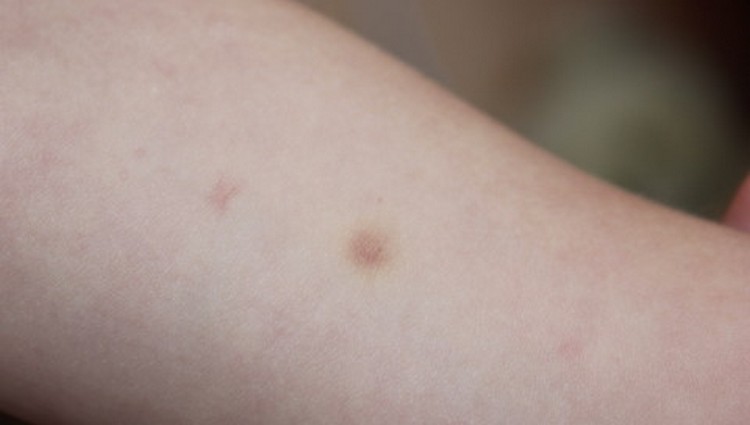 Таким образом, диаскинтест, ти-спот и другие тесты позволяют определить, болен ли человек туберкулезом вообще (легочным или внелегочным), а ПЦР — болен ли он им в активной форме.
Таким образом, диаскинтест, ти-спот и другие тесты позволяют определить, болен ли человек туберкулезом вообще (легочным или внелегочным), а ПЦР — болен ли он им в активной форме.
Флюорография
Всем взрослым с 15 лет в России рекомендуется раз в год проходить флюорографию. Детям с 15 до 17 лет разрешено выполнять Флюрографию, но это не отменяет постановку Диаскинтеста. Справку со свежим снимком требуют везде — даже при посещении ребенка в больнице. Флюорография — это скорее «коллективный барьер» на пути туберкулеза, чем метод выбора для индивидуальной проверки. Флюрография выявляет только легочные формы туберкулеза.
Если у вас остались вопросы по методам диагностики туберкулеза — позвоните нам: +7 812 331 00 00, мы ответим на них!
Читайте также:
Проба Манту и ее интерпретация
Indian Dermatol Online J. 2012, январь-апрель; 3 (1): 2–6.
Surajit Nayak
Отделение кожи и венерической болезни, Медицинский колледж и больница MKCG, Берхампур, Орисса, Индия
Басанти Ахарджья
Отделение кожи и венерической болезни, Медицинский колледж и больница MKCG, Берхампур, Орисса, Индия
Отделение Skin and VD, Медицинский колледж и больница MKCG, Берхампур, Орисса, Индия
Адрес для корреспонденции: Dr.Это статья в открытом доступе, распространяемая на условиях Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, что разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинала.
Эта статья цитируется в других статьях в PMC.Abstract
Туберкулиновая кожная проба — одно из немногих исследований, датируемых 19 годами и до сих пор широко используемыми в качестве важного теста для диагностики туберкулеза.Хотя его очень часто используют врачи во всем мире, его интерпретация всегда остается сложной и противоречивой. На его исход, а также на его интерпретацию влияют различные факторы, такие как возраст, иммунологический статус, сопутствующее заболевание и т. Д. Требуется особая осторожность при интерпретации результата и вынесении заключения.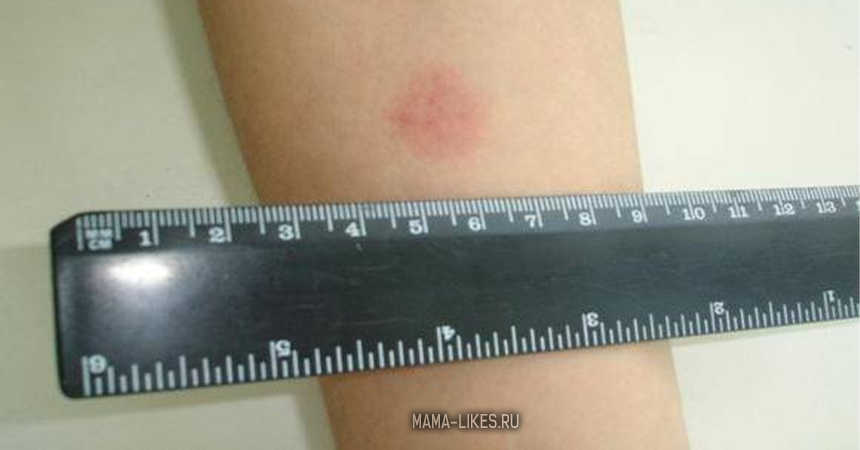 Эта статья была написана с целью разъяснения эффективности и интерпретации стандартного туберкулинового теста.
Эта статья была написана с целью разъяснения эффективности и интерпретации стандартного туберкулинового теста.
Ключевые слова: Интерпретация, проба Манту, туберкулез
ВВЕДЕНИЕ
Туберкулез (ТБ) остается ведущей причиной заболеваемости и смертности в мире, особенно в развивающихся странах.Сочетание факторов, включая высокую стоимость, ограниченные ресурсы и низкую эффективность различных диагностических тестов, затрудняет диагностику ТБ в развивающихся странах. За исключением демонстрации жизнеспособных организмов в тканях и жидкостях организма, туберкулиновая кожная проба (ТКП) является единственным методом выявления инфекции M. tuberculosis у человека и используется для диагностики ТБ у отдельных пациентов, а также в эпидемиологических условиях. , чтобы измерить распространенность туберкулезной инфекции среди населения.
Он был разработан Кохом в 1890 году, но внутрикожная техника, используемая в настоящее время, была описана в 1912 году Шарлем Манту, французским врачом, который разработал свой тест в 1907 году на основе работ Коха и Клеменса фон Пиркетто [1]. длинная история удивительно, что интерпретация теста остается спорной.
длинная история удивительно, что интерпретация теста остается спорной.
Однако различные факторы, присущие как хозяину, так и самому тесту, снижают его специфичность и чувствительность. Следовательно, его применение в любой группе пациентов обычно дает широкий спектр результатов — от наличия реакции у неинфицированных детей до полного отсутствия реакции у некоторых детей с подтвержденным туберкулезом.Распределение результатов обычно подпадает под одну из двух схем в зависимости от количества ложноположительных результатов (перекрестных реакций от других микобактериальных инфекций) в популяции.
Наиболее широко используемым туберкулином является очищенное производное белка (PPD), полученное из культур M. tuberculosis . «Старый туберкулин» для этой цели больше не используется; вместо этого используется более стандартизованный продукт под названием PPD-S (очищенное производное белка, полученное по методу, описанному Зибертом, из M.tuberculosis . PPD нетуберкулезных (т.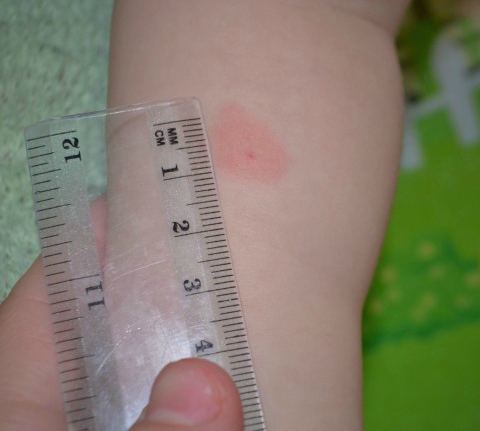 е. атипичных) микобактерий идентифицируются буквой, отличной от S. PPD-A происходит от M. avium ; PPD-G из штамма шотохромогена Gause; PPD-B из нефотохромогенных бацилл Бэтти; PPD-F из быстрорастущего растения M. fortuitum и PPD-Y из желтого фотохромогена M. kanasasii . Это PPD-RT (исследовательский туберкулин) 23, который широко используется индийскими клиницистами, его нелегко получить и могут иметь другие параметры интерпретации.
е. атипичных) микобактерий идентифицируются буквой, отличной от S. PPD-A происходит от M. avium ; PPD-G из штамма шотохромогена Gause; PPD-B из нефотохромогенных бацилл Бэтти; PPD-F из быстрорастущего растения M. fortuitum и PPD-Y из желтого фотохромогена M. kanasasii . Это PPD-RT (исследовательский туберкулин) 23, который широко используется индийскими клиницистами, его нелегко получить и могут иметь другие параметры интерпретации.
ИММУНОЛОГИЧЕСКАЯ ОСНОВА РЕАКЦИИ ТУБЕРКУЛИНА
Реакция на туберкулин, введенный внутрикожно, является классическим примером реакции гиперчувствительности замедленного типа (клеточного типа). Т-клетки, сенсибилизированные предшествующей инфекцией, привлекаются к участку кожи, где они выделяют лимфокины. Эти лимфокины вызывают уплотнение через местное расширение сосудов, отек, отложение фибрина и привлечение других воспалительных клеток в эту область. Особенности реакции включают ( 1 ) ее замедленное течение, достигающее пика более чем через 24 ч после инъекции антигена; ( 2 ) его затвердевший характер; и ( 3 ) его случайные везикуляции и некрозы.
АДМИНИСТРАЦИЯ
Стандартная доза в пять единиц туберкулина (TU) (0,1 мл) вводится внутрикожно (в кожу) и считывается через 48-72 часа. PPD-RT 23 с твином 80 с концентрацией 1 TU и 2 TU — это стандартизированные туберкулины, доступные в Индии, поставляемые Лабораторией вакцины Bacillus Calmette-Guérin (BCG), Guindy, Chennai. Другие доступные на рынке туберкулины не стандартизированы. Твин 80 — это детергент, добавляемый в туберкулин для предотвращения его адсорбции на стеклянной или пластиковой поверхности.В настоящее время все туберкулины производятся и стандартизируются с использованием Tween 80.
Ожидается, что человек, подвергшийся воздействию бактерий, вызовет иммунный ответ в коже, содержащей бактериальные белки. В целях стандартизации чтения и интерпретации результатов почти повсеместно используется 5 ЕД туберкулина PPD RT23. Его следует вводить строго внутрикожно, используя иглу № 28 или 26 и туберкулиновый шприц, из которого можно точно ввести 0,1 мл. При правильном введении инъекции должно появиться дискретное бледное возвышение кожи (волдырь) диаметром от 6 до 10 мм.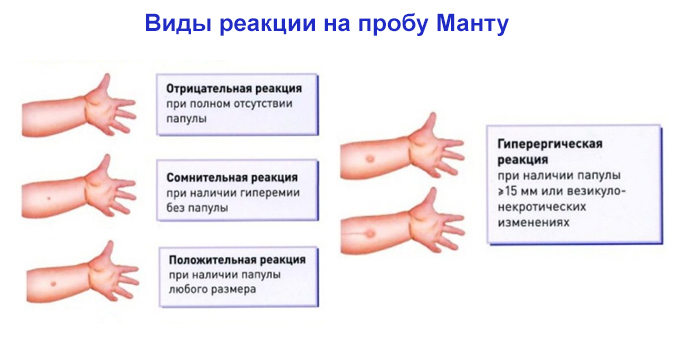 Если будет установлено, что первый тест был проведен неправильно, можно сразу ввести еще одну тестовую дозу, выбрав место в нескольких сантиметрах от первоначальной инъекции. В записи в протоколе должно быть указано место, выбранное для второго теста.
Если будет установлено, что первый тест был проведен неправильно, можно сразу ввести еще одну тестовую дозу, выбрав место в нескольких сантиметрах от первоначальной инъекции. В записи в протоколе должно быть указано место, выбранное для второго теста.
Обычно тест проводится на левом предплечье, чтобы избежать ошибок при чтении. Однако правая рука может использоваться в случае каких-либо противопоказаний к использованию левой руки. Лучше всего проводить тест на ладонной части предплечья. После кожной пробы на туберкулез чрезвычайно важно убедиться, что пациент записался на прием, чтобы прочитать реакцию теста.Пациенту рекомендуется содержать место проведения теста в чистоте, не накрывать его, не царапать и не тереть его. Кожную пробу Манту следует читать между 48 и 72 часами после приема. Чтение следует выполнять при хорошем освещении, слегка согнув предплечье в локтях. Основанием для считывания является наличие или отсутствие уплотнения, которое может быть определено путем осмотра (при виде сбоку против света, а также при прямом освещении) и пальпации.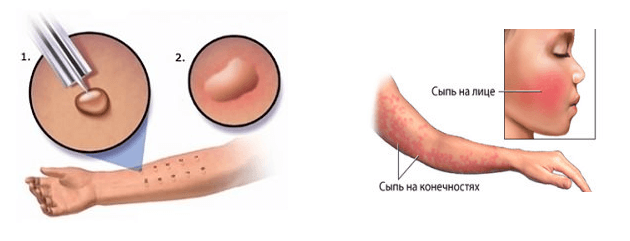 Для стандартизации диаметр уплотнения следует измерять поперек длинной оси предплечья и записывать в миллиметрах.[2,3] Надежное чтение кожной пробы Манту требует стандартизации процедур, обучения, наблюдения и практики. Это может также включать периодические стандартизированные испытания надежности. Следует записать точное измерение уплотнения в миллиметрах (мм). Эритему (покраснение) измерять не следует.
Для стандартизации диаметр уплотнения следует измерять поперек длинной оси предплечья и записывать в миллиметрах.[2,3] Надежное чтение кожной пробы Манту требует стандартизации процедур, обучения, наблюдения и практики. Это может также включать периодические стандартизированные испытания надежности. Следует записать точное измерение уплотнения в миллиметрах (мм). Эритему (покраснение) измерять не следует.
ПОБОЧНОЕ ДЕЙСТВИЕ
Хотя и редко, но были сообщения об анафилактической реакции [4] и реакции на инородное тело [5] на участке пробы Манту. Существует очень небольшой риск серьезной реакции на тест, включая отек и покраснение руки, особенно у людей, которые уже болели туберкулезом или были инфицированы ранее, а также у тех, кто ранее получил вакцину БЦЖ.Аллергические реакции также являются редкими осложнениями. Живые бактерии не используются в тесте, поэтому вероятность развития туберкулеза в результате теста отсутствует. В редких случаях могут возникать такие местные реакции, как регионарный лимфангит и аденит.
ИНТЕРПРЕТАЦИЯ РЕАКЦИИ ТУБЕРКУЛИНА
Проба Манту измеряет не иммунитет к туберкулезу, а степень гиперчувствительности к туберкулину. Нет корреляции между размером уплотнения и вероятностью текущего активного туберкулеза, но размер реакции коррелирует с будущим риском развития туберкулеза.Тест имеет низкую положительную прогностическую ценность для текущего активного заболевания. [6] Нет никакой корреляции между размером поствакцинальных реакций Манту и защитой от туберкулеза, и рутинное тестирование Манту после вакцинации БЦЖ не имеет смысла.
Результаты этого теста следует интерпретировать осторожно. Факторы медицинского риска человека определяют размер уплотнения, результат положительный (5 мм, 10 мм или 15 мм).
Также необходимо сделать запись об образовании пузырьков, пузырей, лимфангита, изъязвлений и некроза на участке исследования.Образование пузырьков, пузырей или некроза на участке исследования указывает на высокую степень чувствительности к туберкулину и, следовательно, на наличие инфекции туберкулезными палочками.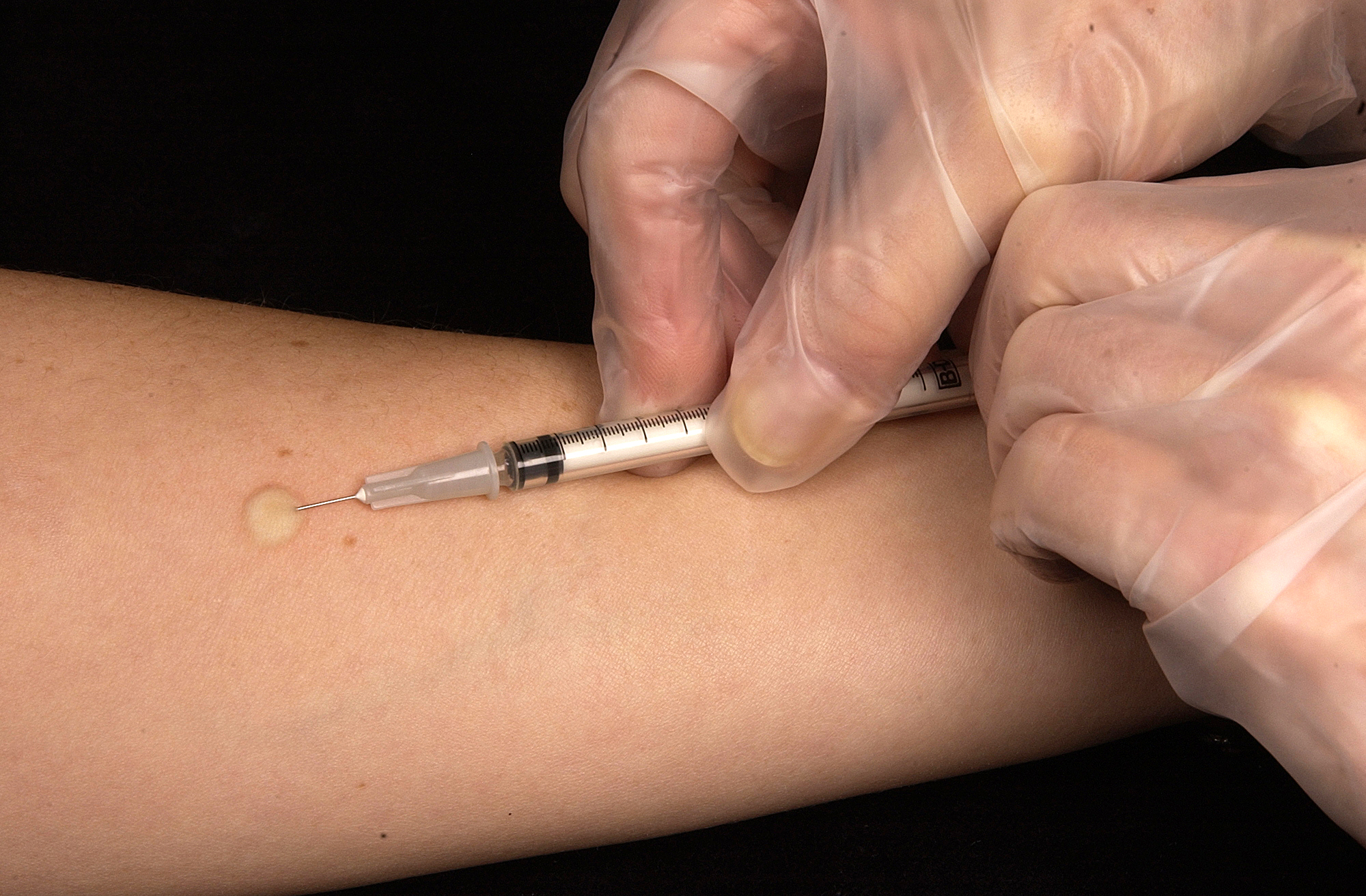 [7]
[7]
Пять мм и более — положительный результат у
ВИЧ-положительный человек
Недавние контакты с активным туберкулезом
Лица с узловыми или фиброзными изменениями на рентгеновском снимке грудной клетки, соответствующими давно вылеченному ТБ
Реципиенты трансплантата органов и другие пациенты с ослабленным иммунитетом, которые принимают цитотоксические иммунодепрессанты, такие как циклофосфамид или метотрексат.
Пациенты, получающие длительную системную кортикостероидную терапию (> шести недель), а также пациенты, принимающие преднизолон в дозе ≥ 15 мг / день или эквивалентной.
Терминальная стадия почечной недостаточности
Десять мм или более положительны в
Недавно прибывшие (менее пяти лет) из стран с высокой распространенностью
Потребители инъекционных наркотиков
Резиденты и сотрудники мест скопления повышенного риска (например,(например, тюрьмы, дома престарелых, больницы, приюты для бездомных и т.
 д.)
д.)Персонал микобактериологической лаборатории
Лица с клиническими состояниями, которые подвергают их высокому риску (например, диабет, длительная терапия кортикостероидами, лейкемия, конечный стадия почечной недостаточности, синдромы хронической мальабсорбции, низкая масса тела и т. д.)
Дети в возрасте до четырех лет или дети и подростки, контактирующие со взрослыми из категорий высокого риска
Младенцы, дети и подростки, подвергшиеся воздействию для взрослых из категорий высокого риска
Пятнадцать мм и более положительны в
Ложноположительный результат
Некоторые люди могут реагировать на ТКП, даже если они не инфицированы M.Туберкулез . Причины этих ложноположительных реакций могут включать в себя, помимо прочего, следующее:
Инфекция нетуберкулезными микобактериями
Предыдущая вакцинация БЦЖ
Неправильный метод проведения TST
Неправильный интерпретация реакции
Использован неправильный флакон с антигеном
Из-за низкой специфичности теста большинство положительных реакций у лиц с низким риском являются ложноположительными. [8] Ложноположительный результат может быть вызван нетуберкулезными микобактериями или предыдущим введением вакцины БЦЖ. Предыдущая вакцинация БЦЖ может привести к ложноположительному результату в течение многих лет [9].
[8] Ложноположительный результат может быть вызван нетуберкулезными микобактериями или предыдущим введением вакцины БЦЖ. Предыдущая вакцинация БЦЖ может привести к ложноположительному результату в течение многих лет [9].
Ложноотрицательный результат
Отрицательный результат Манту обычно означает, что человек никогда не подвергался воздействию M. tuberculosis . Однако есть факторы, которые могут вызвать ложноотрицательный результат или снижение способности реагировать на туберкулин. [10,11]
Кожная анергия (анергия — это неспособность реагировать на кожные пробы из-за ослабленной иммунной системы)
Недавняя инфекция ТБ (в течение 8-10 недель после заражения)
Очень старая инфекция ТБ (много лет)
Очень молодой возраст (менее шести месяцев)
Недавняя вакцинация живым вирусом (е.ж., корь и оспа)
Одностороннее заболевание туберкулезом
Некоторые вирусные заболевания (например, корь и ветряная оспа)
Неправильный метод проведения ТКП
Неправильная интерпретация реакции, недостаточная доза и непреднамеренная подкожная инъекция.
Отсутствие клеточного иммунитета к туберкулину может быть связано с отсутствием предшествующей сенсибилизации или с ложноотрицательным результатом по разным причинам или с анергией из-за подавления иммунитета.Большинство детей с отрицательным результатом не были инфицированы M. tuberculosis . Небольшая часть в остальном нормальных детей с инфекцией M. tuberculosis остается PPD-отрицательной по неизвестным причинам. С момента заражения до развития CMI существует период окна продолжительностью от двух до шести недель, когда проба Манту будет отрицательной. Пациенты с ослабленным иммунитетом, особенно с ВИЧ и низким числом Т-лимфоцитов CD4, часто показывают отрицательные результаты теста PPD.Это связано с тем, что иммунная система должна быть функциональной, чтобы реагировать на производное белка, вводимое под кожу.
Отрицательные тесты можно интерпретировать как то, что человек не был инфицирован бактериями ТБ или что человек был инфицирован недавно, и прошло недостаточно времени, чтобы организм отреагировал на кожный тест. Повторный тест не рекомендуется ранее, чем через неделю, поскольку туберкулин, вводимый для первого теста, оказывает бустер-эффект на последующую дозу. TST может перейти в положительный результат ≤ восьми недель после заражения Mycobacterium tuberculosis , этот интервал обычно называют «периодом окна».Отрицательный результат ТКП, полученный менее чем за восемь недель до этого, не исключает инфекции, и второй тест рекомендуется через восемь недель. [12] Кроме того, поскольку развитие реакции у пожилого человека может занять более 72 часов, может оказаться полезным повторить кожную пробу на ТБ через 96 часов и еще раз через одну неделю для адекватного скрининга этих людей. Люди с ослабленным иммунитетом могут быть не в состоянии адекватно отреагировать на пробу Манту, и может потребоваться либо рентген грудной клетки, либо образец мокроты.
Повторный тест не рекомендуется ранее, чем через неделю, поскольку туберкулин, вводимый для первого теста, оказывает бустер-эффект на последующую дозу. TST может перейти в положительный результат ≤ восьми недель после заражения Mycobacterium tuberculosis , этот интервал обычно называют «периодом окна».Отрицательный результат ТКП, полученный менее чем за восемь недель до этого, не исключает инфекции, и второй тест рекомендуется через восемь недель. [12] Кроме того, поскольку развитие реакции у пожилого человека может занять более 72 часов, может оказаться полезным повторить кожную пробу на ТБ через 96 часов и еще раз через одну неделю для адекватного скрининга этих людей. Люди с ослабленным иммунитетом могут быть не в состоянии адекватно отреагировать на пробу Манту, и может потребоваться либо рентген грудной клетки, либо образец мокроты.
Интерпретация для детей: Правильно проведенная проба Манту может иметь неоценимое значение при обследовании ребенка с подозрением на ТБ.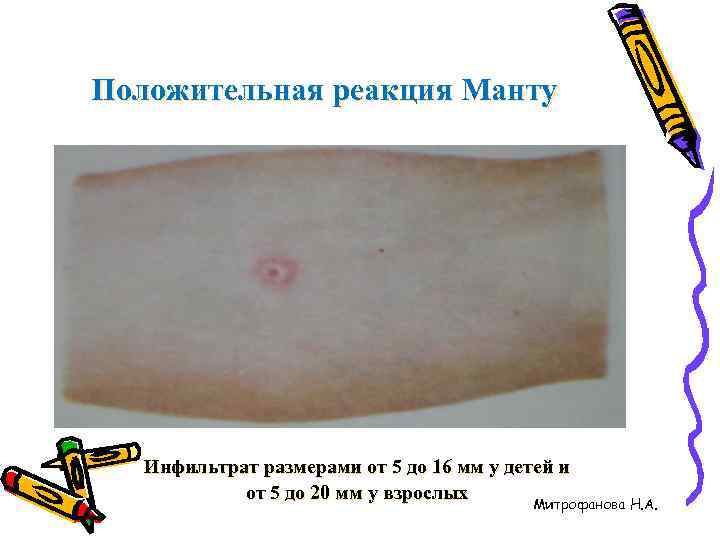 Однако интерпретация результата часто затруднена, так как разные работники используют уплотнения разного размера, чтобы указать на положительную реакцию. Хотя сам тест не является ни на 100% ни чувствительным, ни на 100% специфичным, прогностическая ценность положительной реакции очень высока в такой группе. Ранее было показано, что недоедание влияет на результаты туберкулиновой пробы. Как и в других исследованиях, у детей с недостаточным весом в этом исследовании вероятность получения отрицательного результата по Манту была значительно выше [].
Однако интерпретация результата часто затруднена, так как разные работники используют уплотнения разного размера, чтобы указать на положительную реакцию. Хотя сам тест не является ни на 100% ни чувствительным, ни на 100% специфичным, прогностическая ценность положительной реакции очень высока в такой группе. Ранее было показано, что недоедание влияет на результаты туберкулиновой пробы. Как и в других исследованиях, у детей с недостаточным весом в этом исследовании вероятность получения отрицательного результата по Манту была значительно выше [].
Таблица 1
Пороговая величина реакции при положительном результате пробы Манту у детей
Как и во многих других исследованиях, у большинства детей не было реакции на туберкулин, несмотря на то, что они получили иммунизацию БЦЖ вскоре после рождения. Причины этого не всегда ясны, но ясно, что любая чувствительность к туберкулину, которую могла вызвать БЦЖ, не могла быть значительной или устойчивой. Это согласуется с нынешней рекомендацией о том, что для пациентов с высоким риском туберкулеза история вакцинации БЦЖ не должна учитываться при интерпретации туберкулиновой пробы.
УСИЛИТЕЛЬНЫЙ ЭФФЕКТ
У некоторых людей, инфицированных M. tuberculosis , способность реагировать на туберкулин со временем может уменьшаться. При проведении ТКП через несколько лет после заражения у этих людей может быть ложноотрицательная реакция. Однако TST может стимулировать иммунную систему, вызывая положительную или повышенную реакцию на последующие тесты. Проведение второй TST после первоначальной отрицательной реакции TST называется двухэтапным тестированием. Когда сенсибилизация к микобактериям произошла много лет назад, первоначальная внутрикожная инъекция туберкулина может вызвать отрицательный или слабоположительный ответ из-за того, что в кровотоке слишком мало сенсибилизированных лимфоцитов, чтобы вызвать значительный местный ответ.Если тест повторяется, большее значение может быть получено из-за того, что иммунный ответ «отзывается» или «усиливается» первым тестом. Второе усиленное показание является правильным, то есть результатом, который следует использовать для принятия решения или будущего сравнения. Повышение является максимальным, если второй тест проводится между одной и пятью неделями после первоначального теста, и он может продолжаться до двух лет.
Повышение является максимальным, если второй тест проводится между одной и пятью неделями после первоначального теста, и он может продолжаться до двух лет.
РЕВЕРСИЯ МАНТУ
Реверсирование определяется как изменение отрицательного результата Манту после предыдущего положительного результата.Как правило, это явление не характерно для здоровых людей и встречается менее чем у 10% таких людей с ранее положительным диагнозом Манту.
Реверсия более распространена [13]
у пожилых людей (по оценкам, 8% в год)
, когда исходное значение Манту <14 мм
у тех, у кого исходная положительная реакция была усилена результат (определяется двухэтапным тестированием).
КОНВЕРСИЯ МАНТУ
В то время как бустинг — это отзыв реакции гиперчувствительности в отсутствие новой инфекции, конверсия — это развитие новой или повышенной гиперчувствительности из-за инфекции туберкулезными или нетуберкулезными микобактериями, включая вакцинацию БЦЖ.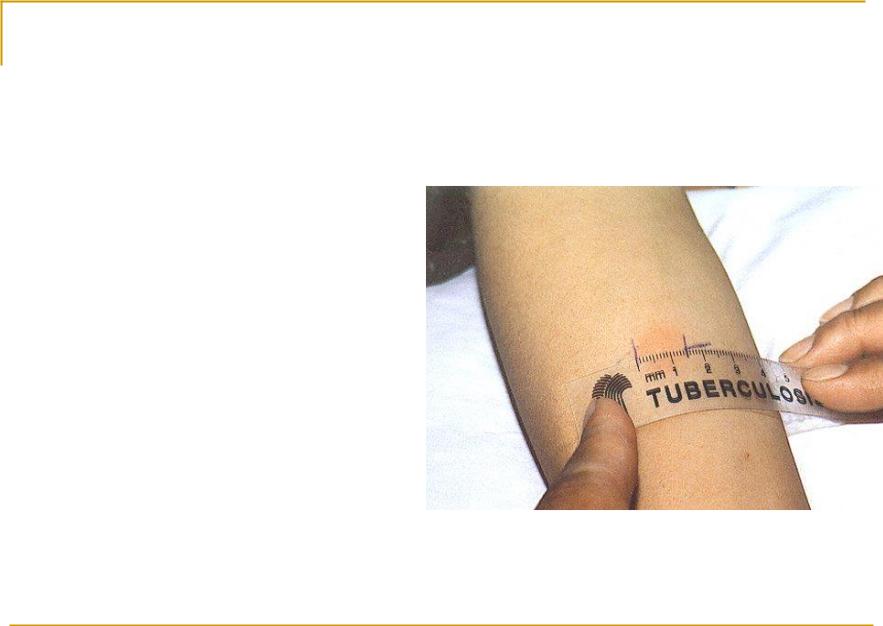
Конверсия Манту определяется [14] как изменение (в течение двухлетнего периода) реактивности Манту, которое соответствует любому из следующих критериев:
изменение с отрицательной реакции на положительную
увеличение ≥ 10 мм.
Конверсия была связана с ежегодной заболеваемостью туберкулезом 4% среди подростков [15] или 6% среди лиц, контактировавших с пациентами с положительным мазком [16].
Споры ведутся о времени, необходимом для иммунологических изменений, вызывающих конверсию Манту после инфекции.После непреднамеренной вакцинации M. tuberculosis (катастрофа в Любеке) у детей развились положительные реакции в течение трех-семи недель. Другие исследования показали клиническое заболевание с положительным туберкулиновым тестом в период от 19 до 57 дней после заражения, в среднем 37 дней. [1]
Следовательно, при проверке контактных лиц на туберкулез на предмет конверсии второй туберкулиновый тест проводится через восемь недель после даты последнего контакта с больным-источником. (В прошлом использовался традиционный период окна или интервал в 12 недель.)
(В прошлом использовался традиционный период окна или интервал в 12 недель.)
ВАКЦИНА КАЛЬМЕТТА-ГЕРИНА И ТЕСТ МАНТУ
Существуют разногласия относительно роли теста Манту у вакцинированных людей. Согласно рекомендациям США, ТКП не противопоказана людям, вакцинированным БЦЖ, и что предшествующая вакцинация БЦЖ не должна влиять на интерпретацию теста.
В соответствии с руководящими принципами США диагностика и лечение латентной туберкулезной инфекции (ЛТБИ) и лечение ЛТИ рассматриваются для любого вакцинированного БЦЖ человека, чья кожная проба составляет 10 мм или больше, при наличии любого из этих обстоятельств:
Был в контакте с другим человеком, больным инфекционным ТБ
Родился или жил в стране с высокой распространенностью ТБ
Постоянно контактирует с группами населения с высокой распространенностью ТБ.
СИТУАЦИИ, КОТОРЫЕ НЕ РЕКОМЕНДУЕТСЯ ТЕСТИРОВАНИЕ МАНТУ
Тест Манту не рекомендуется в следующих ситуациях:
Прошлые реакции Манту ≥ 15 мм: повторение теста не даст новой диагностической информации и создаст дискомфорт
Перенесенное заболевание туберкулезом: не будет получена полезная диагностическая информация и вероятен значительный дискомфорт
Младенцы до 12 недель: положительная реакция очень важна, но отрицательная реакция может указывать на то, что ребенок слишком мал, чтобы дать ответ , и тест необходимо будет повторить, если облучение произошло.
 Тестирование Манту перед вакцинацией до достижения 12-недельного возраста не требуется, если ребенок не инфицирован туберкулезом.
Тестирование Манту перед вакцинацией до достижения 12-недельного возраста не требуется, если ребенок не инфицирован туберкулезом.
ПОСЛЕДНИЕ РАЗРАБОТКИ
Тест Манту технически сложен для проведения и чтения, поэтому могут возникнуть ложные показания, если тестировщик не обладает достаточными навыками. Если проводится двухэтапный тест, может потребоваться четыре посещения пациента, а соблюдение этого условия иногда затруднено. Было бы проще сделать анализ, который можно сделать во время одного посещения пациента, например анализ крови.
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов FDA одобрило новый диагностический тест (QuantiFERON-TB GOLD, производимый Cellestis, Inc.) на туберкулез. Анализ крови выявляет наличие инфекции Mycobacterium tuberculosis (TB) путем измерения интерферона-гамма (IFN-G), собранного в плазме цельной крови, инкубированной со специфическими антигенами M. tuberculosis , ESAT-6 (QFT-RD1 ) и CFP-10. Этот новый иммунодиагностический тест был запущен в качестве вспомогательного средства в диагностике ЛТИ. Комбинация ESAT-6 и CFP10 оказалась высокочувствительной и специфичной как для диагностики in vivo, и in vitro, так и для диагностики .У людей комбинация имела высокую чувствительность (73%) и гораздо более высокую специфичность (93%), чем PPD (7%). Тест QFT-RD1 чувствителен для диагностики ТБ, особенно у пациентов с отрицательными результатами микроскопии и посева. Несмотря на то, что антигены, такие как ESAT-6 и CFP10, не ограничиваются M. tuberculosis , они обещают специфическое обнаружение инфекции M. tuberculosis и могут быть очень полезным дополнительным инструментом для диагностики туберкулеза. .
Этот новый иммунодиагностический тест был запущен в качестве вспомогательного средства в диагностике ЛТИ. Комбинация ESAT-6 и CFP10 оказалась высокочувствительной и специфичной как для диагностики in vivo, и in vitro, так и для диагностики .У людей комбинация имела высокую чувствительность (73%) и гораздо более высокую специфичность (93%), чем PPD (7%). Тест QFT-RD1 чувствителен для диагностики ТБ, особенно у пациентов с отрицательными результатами микроскопии и посева. Несмотря на то, что антигены, такие как ESAT-6 и CFP10, не ограничиваются M. tuberculosis , они обещают специфическое обнаружение инфекции M. tuberculosis и могут быть очень полезным дополнительным инструментом для диагностики туберкулеза. .
Рекомендации по использованию теста QuantiFERON были выпущены [Центром контроля заболеваний] в декабре 2005 года.В настоящее время позиция КвантиФЕРОН-ТБ в диагностике ЛТИ не ясна. Возможно, в будущем появится возможность заменить кожную пробу этим или альтернативным анализом in vitro .
Сноски
Источник поддержки: Нет
Конфликт интересов: Не объявлен.
СПИСОК ЛИТЕРАТУРЫ
1. Мензис Д. Туберкулиновая кожная проба. В: Райхман Л.Б., Хершфилд Е.С., редакторы. Туберкулез: комплексный международный подход. Нью-Йорк: Марсель Деккер; 2000 г.С. 279–322. [Google Scholar] 2. Ховард А., Мерсер П., Натарадж ХК, Канг BC. Скос вниз лучше скоса вверх при внутрикожном кожном тестировании. Ann Allergy Asthma Immunol. 1997. 78: 594–6. [PubMed] [Google Scholar] 3. Американское торакальное общество. Кожная туберкулиновая проба, 1981. Am Rev Respir Dis. 1981; 124: 346–51. [Google Scholar] 4. Риск серьезных аллергических реакций после введения туберсола [очищенного производного белка туберкулина (Манту)] [письмо уважаемому специалисту в области здравоохранения] Оттава: Министерство здравоохранения Канады; 2005 г.Санофи Пастер Лимитед, Отделение продуктов здравоохранения и продуктов здравоохранения Министерства здравоохранения Канады, Агентство общественного здравоохранения Канады. [Google Scholar] 5. Ким Й., Доус-Хиггс Э., Загарелла С. Реакция на инородное тело с участком пробы Манту. Australas J Dermatol. 2005. 46: 169–71. [PubMed] [Google Scholar] 6. Аль Захрани К., Аль Джахдали Х., Мензис Д. Имеет ли значение размер? Полезность величины туберкулиновых реакций для диагностики микобактериальных заболеваний. Am J Resp Crit Care Med. 2000. 162: 1419–22. [PubMed] [Google Scholar] 7. Американское торакальное общество.Заявление о туберкулиновой кожной пробе Американского торакального общества, Медицинской секции Американской ассоциации легких. Am Rev Respir Dis. 1981; 124: 356–63. [Google Scholar] 8. Старке-младший. Кожные пробы на туберкулез: новые школы мысли. J Am Acad Pediatr. 1996; 98: 123–5. [PubMed] [Google Scholar] 9. Чатурведи Н., Кокрофт А. Скрининг на туберкулез среди медицинских работников: кому нужны рентгеновские снимки грудной клетки? Оккуп Мед (Лондон) 1992; 42: 179–82. [PubMed] [Google Scholar] 10. Американское торакальное общество. Стандарты диагностики и классификация туберкулеза у взрослых и детей.
[Google Scholar] 5. Ким Й., Доус-Хиггс Э., Загарелла С. Реакция на инородное тело с участком пробы Манту. Australas J Dermatol. 2005. 46: 169–71. [PubMed] [Google Scholar] 6. Аль Захрани К., Аль Джахдали Х., Мензис Д. Имеет ли значение размер? Полезность величины туберкулиновых реакций для диагностики микобактериальных заболеваний. Am J Resp Crit Care Med. 2000. 162: 1419–22. [PubMed] [Google Scholar] 7. Американское торакальное общество.Заявление о туберкулиновой кожной пробе Американского торакального общества, Медицинской секции Американской ассоциации легких. Am Rev Respir Dis. 1981; 124: 356–63. [Google Scholar] 8. Старке-младший. Кожные пробы на туберкулез: новые школы мысли. J Am Acad Pediatr. 1996; 98: 123–5. [PubMed] [Google Scholar] 9. Чатурведи Н., Кокрофт А. Скрининг на туберкулез среди медицинских работников: кому нужны рентгеновские снимки грудной клетки? Оккуп Мед (Лондон) 1992; 42: 179–82. [PubMed] [Google Scholar] 10. Американское торакальное общество. Стандарты диагностики и классификация туберкулеза у взрослых и детей.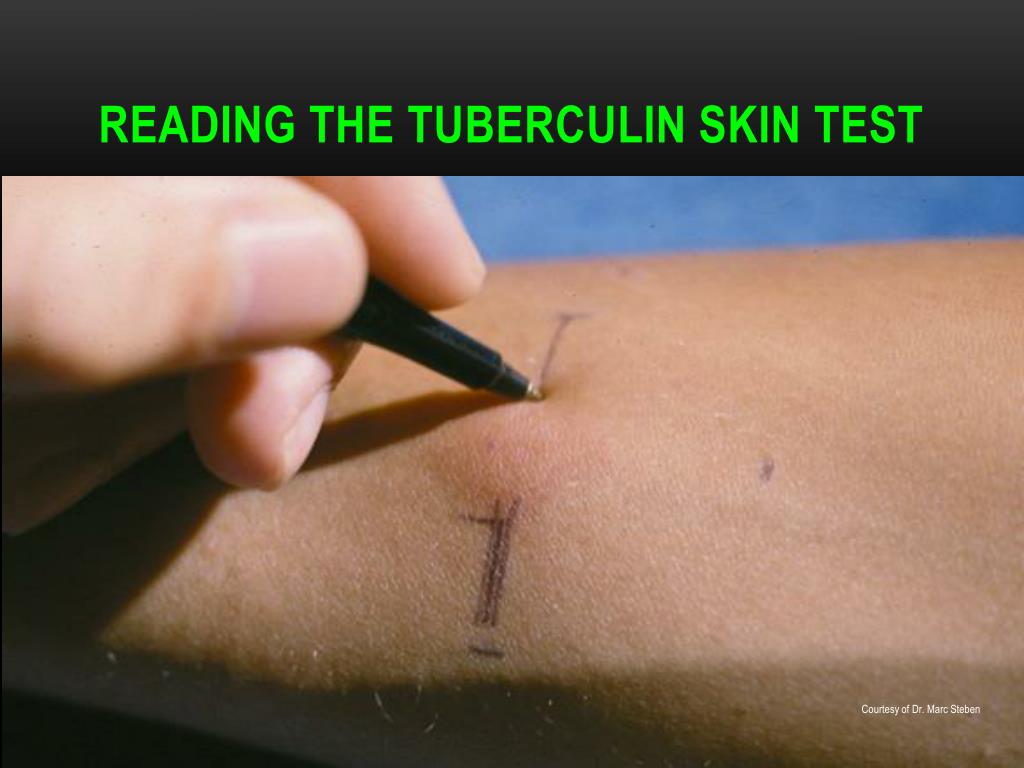 Am J Respir Crit Care Med. 2000; 161: 1376–95. [PubMed] [Google Scholar] 11. Американское торакальное общество / Центры контроля заболеваний. Целевое туберкулиновое тестирование и лечение скрытой туберкулезной инфекции. Am J Respir Crit Care Med. 2000; 161: S221–47. [PubMed] [Google Scholar] 12. Anibarro L, Trigo M, Villaverde C, Pena A, Cortizo S, Sande D и др. Анализ высвобождения интерферона-γ у лиц, контактировавших с туберкулезом: есть ли период окна? Eur Respir J. 2011; 37: 215–7. [PubMed] [Google Scholar] 13. Мензис Д. Интерпретация повторных туберкулиновых проб.Повышение, преобразование и возврат. Am J Respir Crit Care Med. 1999; 159: 15–21. [PubMed] [Google Scholar] 14. Целевое туберкулиновое тестирование и лечение скрытой туберкулезной инфекции. Американское торакальное общество. MMWR Recomm Rep. 2000; 49: 1–51. [PubMed] [Google Scholar] 15. Сазерленд И. Эволюция клинического туберкулеза у подростков. Бугорок. 1966; 47: 308. [Google Scholar] 16. Veening GJ. Долгосрочная профилактика изониазидом.
Am J Respir Crit Care Med. 2000; 161: 1376–95. [PubMed] [Google Scholar] 11. Американское торакальное общество / Центры контроля заболеваний. Целевое туберкулиновое тестирование и лечение скрытой туберкулезной инфекции. Am J Respir Crit Care Med. 2000; 161: S221–47. [PubMed] [Google Scholar] 12. Anibarro L, Trigo M, Villaverde C, Pena A, Cortizo S, Sande D и др. Анализ высвобождения интерферона-γ у лиц, контактировавших с туберкулезом: есть ли период окна? Eur Respir J. 2011; 37: 215–7. [PubMed] [Google Scholar] 13. Мензис Д. Интерпретация повторных туберкулиновых проб.Повышение, преобразование и возврат. Am J Respir Crit Care Med. 1999; 159: 15–21. [PubMed] [Google Scholar] 14. Целевое туберкулиновое тестирование и лечение скрытой туберкулезной инфекции. Американское торакальное общество. MMWR Recomm Rep. 2000; 49: 1–51. [PubMed] [Google Scholar] 15. Сазерленд И. Эволюция клинического туберкулеза у подростков. Бугорок. 1966; 47: 308. [Google Scholar] 16. Veening GJ. Долгосрочная профилактика изониазидом. Контролируемое испытание по профилактике изониазидом после недавней конверсии туберкулина у молодых людей.Bull Int Union Tuberc. 1968; 41: 169–71. [PubMed] [Google Scholar]
Контролируемое испытание по профилактике изониазидом после недавней конверсии туберкулина у молодых людей.Bull Int Union Tuberc. 1968; 41: 169–71. [PubMed] [Google Scholar]Тест Манту и его интерпретация
Indian Dermatol Online J. 2012, январь-апрель; 3 (1): 2–6.
Surajit Nayak
Отделение кожи и венерической болезни, Медицинский колледж и больница MKCG, Берхампур, Орисса, Индия
Басанти Ахарджья
Отделение кожи и венерической болезни, Медицинский колледж и больница MKCG, Берхампур, Орисса, Индия
Отделение Skin and VD, Медицинский колледж и больница MKCG, Берхампур, Орисса, Индия
Адрес для корреспонденции: Dr.Сураджит Наяк, кафедра кожи и венерической болезни, Медицинский колледж MKCG, Берхампур, Орисса, Индия. Электронная почта: ni.oc.oohay@kyntijarus Авторские права: © Indian Dermatology Online Journal Это статья в открытом доступе, распространяемая на условиях Creative Commons Attribution-Noncommercial-Share Alike 3. 0 Unported, что разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинала.
0 Unported, что разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинала.
Abstract
Туберкулиновая кожная проба — одно из немногих исследований, датируемых 19 годами и до сих пор широко используемыми в качестве важного теста для диагностики туберкулеза.Хотя его очень часто используют врачи во всем мире, его интерпретация всегда остается сложной и противоречивой. На его исход, а также на его интерпретацию влияют различные факторы, такие как возраст, иммунологический статус, сопутствующее заболевание и т. Д. Требуется особая осторожность при интерпретации результата и вынесении заключения. Эта статья была написана с целью разъяснения эффективности и интерпретации стандартного туберкулинового теста.
Ключевые слова: Интерпретация, проба Манту, туберкулез
ВВЕДЕНИЕ
Туберкулез (ТБ) остается ведущей причиной заболеваемости и смертности в мире, особенно в развивающихся странах.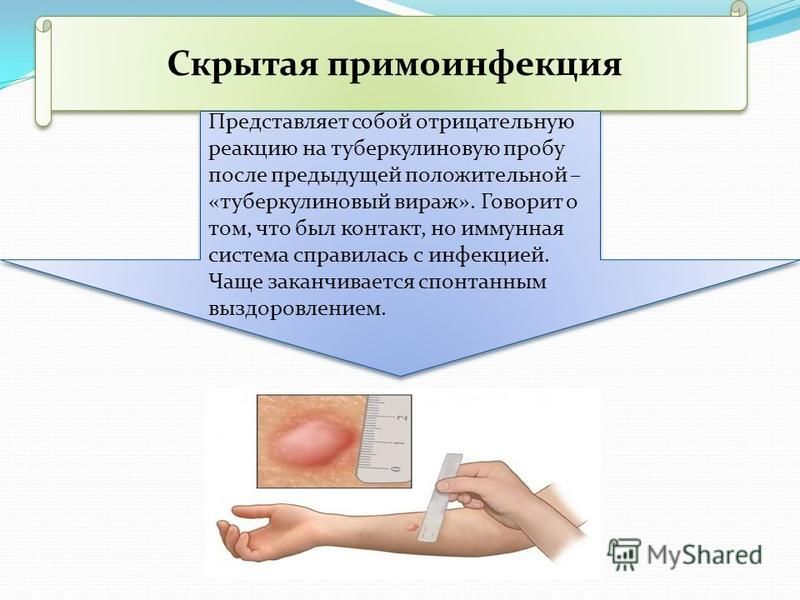 Сочетание факторов, включая высокую стоимость, ограниченные ресурсы и низкую эффективность различных диагностических тестов, затрудняет диагностику ТБ в развивающихся странах. За исключением демонстрации жизнеспособных организмов в тканях и жидкостях организма, туберкулиновая кожная проба (ТКП) является единственным методом выявления инфекции M. tuberculosis у человека и используется для диагностики ТБ у отдельных пациентов, а также в эпидемиологических условиях. , чтобы измерить распространенность туберкулезной инфекции среди населения.
Сочетание факторов, включая высокую стоимость, ограниченные ресурсы и низкую эффективность различных диагностических тестов, затрудняет диагностику ТБ в развивающихся странах. За исключением демонстрации жизнеспособных организмов в тканях и жидкостях организма, туберкулиновая кожная проба (ТКП) является единственным методом выявления инфекции M. tuberculosis у человека и используется для диагностики ТБ у отдельных пациентов, а также в эпидемиологических условиях. , чтобы измерить распространенность туберкулезной инфекции среди населения.
Он был разработан Кохом в 1890 году, но внутрикожная техника, используемая в настоящее время, была описана в 1912 году Шарлем Манту, французским врачом, который разработал свой тест в 1907 году на основе работ Коха и Клеменса фон Пиркетто [1]. длинная история удивительно, что интерпретация теста остается спорной.
Однако различные факторы, присущие как хозяину, так и самому тесту, снижают его специфичность и чувствительность. Следовательно, его применение в любой группе пациентов обычно дает широкий спектр результатов — от наличия реакции у неинфицированных детей до полного отсутствия реакции у некоторых детей с подтвержденным туберкулезом. Распределение результатов обычно подпадает под одну из двух схем в зависимости от количества ложноположительных результатов (перекрестных реакций от других микобактериальных инфекций) в популяции.
Распределение результатов обычно подпадает под одну из двух схем в зависимости от количества ложноположительных результатов (перекрестных реакций от других микобактериальных инфекций) в популяции.
Наиболее широко используемым туберкулином является очищенное производное белка (PPD), полученное из культур M. tuberculosis . «Старый туберкулин» для этой цели больше не используется; вместо этого используется более стандартизованный продукт под названием PPD-S (очищенное производное белка, полученное по методу, описанному Зибертом, из M.tuberculosis . PPD нетуберкулезных (т.е. атипичных) микобактерий идентифицируются буквой, отличной от S. PPD-A происходит от M. avium ; PPD-G из штамма шотохромогена Gause; PPD-B из нефотохромогенных бацилл Бэтти; PPD-F из быстрорастущего растения M. fortuitum и PPD-Y из желтого фотохромогена M. kanasasii . Это PPD-RT (исследовательский туберкулин) 23, который широко используется индийскими клиницистами, его нелегко получить и могут иметь другие параметры интерпретации.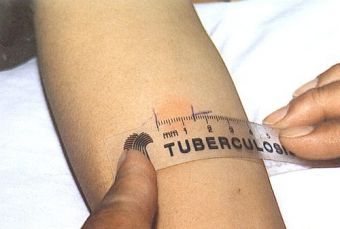
ИММУНОЛОГИЧЕСКАЯ ОСНОВА РЕАКЦИИ ТУБЕРКУЛИНА
Реакция на туберкулин, введенный внутрикожно, является классическим примером реакции гиперчувствительности замедленного типа (клеточного типа). Т-клетки, сенсибилизированные предшествующей инфекцией, привлекаются к участку кожи, где они выделяют лимфокины. Эти лимфокины вызывают уплотнение через местное расширение сосудов, отек, отложение фибрина и привлечение других воспалительных клеток в эту область. Особенности реакции включают ( 1 ) ее замедленное течение, достигающее пика более чем через 24 ч после инъекции антигена; ( 2 ) его затвердевший характер; и ( 3 ) его случайные везикуляции и некрозы.
АДМИНИСТРАЦИЯ
Стандартная доза в пять единиц туберкулина (TU) (0,1 мл) вводится внутрикожно (в кожу) и считывается через 48-72 часа. PPD-RT 23 с твином 80 с концентрацией 1 TU и 2 TU — это стандартизированные туберкулины, доступные в Индии, поставляемые Лабораторией вакцины Bacillus Calmette-Guérin (BCG), Guindy, Chennai. Другие доступные на рынке туберкулины не стандартизированы. Твин 80 — это детергент, добавляемый в туберкулин для предотвращения его адсорбции на стеклянной или пластиковой поверхности.В настоящее время все туберкулины производятся и стандартизируются с использованием Tween 80.
Другие доступные на рынке туберкулины не стандартизированы. Твин 80 — это детергент, добавляемый в туберкулин для предотвращения его адсорбции на стеклянной или пластиковой поверхности.В настоящее время все туберкулины производятся и стандартизируются с использованием Tween 80.
Ожидается, что человек, подвергшийся воздействию бактерий, вызовет иммунный ответ в коже, содержащей бактериальные белки. В целях стандартизации чтения и интерпретации результатов почти повсеместно используется 5 ЕД туберкулина PPD RT23. Его следует вводить строго внутрикожно, используя иглу № 28 или 26 и туберкулиновый шприц, из которого можно точно ввести 0,1 мл. При правильном введении инъекции должно появиться дискретное бледное возвышение кожи (волдырь) диаметром от 6 до 10 мм.Если будет установлено, что первый тест был проведен неправильно, можно сразу ввести еще одну тестовую дозу, выбрав место в нескольких сантиметрах от первоначальной инъекции. В записи в протоколе должно быть указано место, выбранное для второго теста.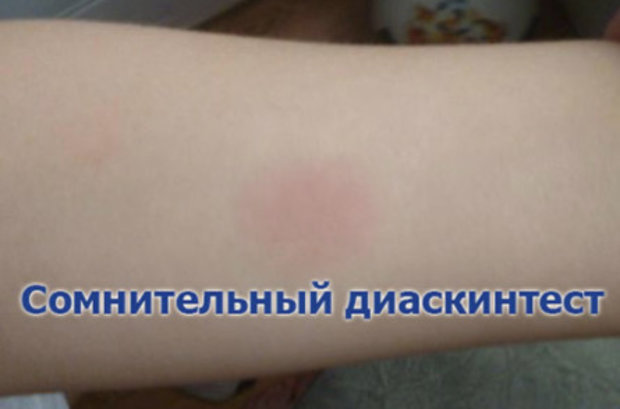
Обычно тест проводится на левом предплечье, чтобы избежать ошибок при чтении. Однако правая рука может использоваться в случае каких-либо противопоказаний к использованию левой руки. Лучше всего проводить тест на ладонной части предплечья. После кожной пробы на туберкулез чрезвычайно важно убедиться, что пациент записался на прием, чтобы прочитать реакцию теста.Пациенту рекомендуется содержать место проведения теста в чистоте, не накрывать его, не царапать и не тереть его. Кожную пробу Манту следует читать между 48 и 72 часами после приема. Чтение следует выполнять при хорошем освещении, слегка согнув предплечье в локтях. Основанием для считывания является наличие или отсутствие уплотнения, которое может быть определено путем осмотра (при виде сбоку против света, а также при прямом освещении) и пальпации. Для стандартизации диаметр уплотнения следует измерять поперек длинной оси предплечья и записывать в миллиметрах.[2,3] Надежное чтение кожной пробы Манту требует стандартизации процедур, обучения, наблюдения и практики. Это может также включать периодические стандартизированные испытания надежности. Следует записать точное измерение уплотнения в миллиметрах (мм). Эритему (покраснение) измерять не следует.
Это может также включать периодические стандартизированные испытания надежности. Следует записать точное измерение уплотнения в миллиметрах (мм). Эритему (покраснение) измерять не следует.
ПОБОЧНОЕ ДЕЙСТВИЕ
Хотя и редко, но были сообщения об анафилактической реакции [4] и реакции на инородное тело [5] на участке пробы Манту. Существует очень небольшой риск серьезной реакции на тест, включая отек и покраснение руки, особенно у людей, которые уже болели туберкулезом или были инфицированы ранее, а также у тех, кто ранее получил вакцину БЦЖ.Аллергические реакции также являются редкими осложнениями. Живые бактерии не используются в тесте, поэтому вероятность развития туберкулеза в результате теста отсутствует. В редких случаях могут возникать такие местные реакции, как регионарный лимфангит и аденит.
ИНТЕРПРЕТАЦИЯ РЕАКЦИИ ТУБЕРКУЛИНА
Проба Манту измеряет не иммунитет к туберкулезу, а степень гиперчувствительности к туберкулину. Нет корреляции между размером уплотнения и вероятностью текущего активного туберкулеза, но размер реакции коррелирует с будущим риском развития туберкулеза. Тест имеет низкую положительную прогностическую ценность для текущего активного заболевания. [6] Нет никакой корреляции между размером поствакцинальных реакций Манту и защитой от туберкулеза, и рутинное тестирование Манту после вакцинации БЦЖ не имеет смысла.
Тест имеет низкую положительную прогностическую ценность для текущего активного заболевания. [6] Нет никакой корреляции между размером поствакцинальных реакций Манту и защитой от туберкулеза, и рутинное тестирование Манту после вакцинации БЦЖ не имеет смысла.
Результаты этого теста следует интерпретировать осторожно. Факторы медицинского риска человека определяют размер уплотнения, результат положительный (5 мм, 10 мм или 15 мм).
Также необходимо сделать запись об образовании пузырьков, пузырей, лимфангита, изъязвлений и некроза на участке исследования.Образование пузырьков, пузырей или некроза на участке исследования указывает на высокую степень чувствительности к туберкулину и, следовательно, на наличие инфекции туберкулезными палочками. [7]
Пять мм и более — положительный результат у
ВИЧ-положительный человек
Недавние контакты с активным туберкулезом
Лица с узловыми или фиброзными изменениями на рентгеновском снимке грудной клетки, соответствующими давно вылеченному ТБ
Реципиенты трансплантата органов и другие пациенты с ослабленным иммунитетом, которые принимают цитотоксические иммунодепрессанты, такие как циклофосфамид или метотрексат.
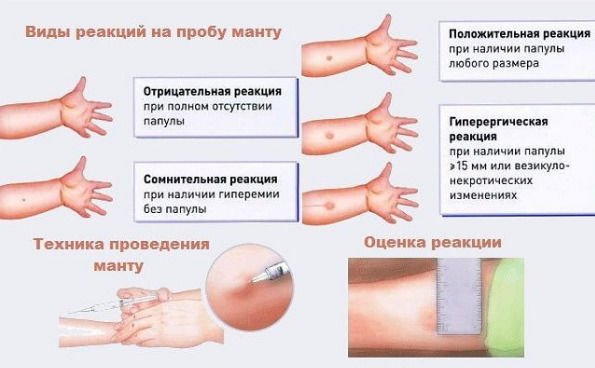
Пациенты, получающие длительную системную кортикостероидную терапию (> шести недель), а также пациенты, принимающие преднизолон в дозе ≥ 15 мг / день или эквивалентной.
Терминальная стадия почечной недостаточности
Десять мм или более положительны в
Недавно прибывшие (менее пяти лет) из стран с высокой распространенностью
Потребители инъекционных наркотиков
Резиденты и сотрудники мест скопления повышенного риска (например,(например, тюрьмы, дома престарелых, больницы, приюты для бездомных и т. д.)
Персонал микобактериологической лаборатории
Лица с клиническими состояниями, которые подвергают их высокому риску (например, диабет, длительная терапия кортикостероидами, лейкемия, конечный стадия почечной недостаточности, синдромы хронической мальабсорбции, низкая масса тела и т. д.)
Дети в возрасте до четырех лет или дети и подростки, контактирующие со взрослыми из категорий высокого риска
Младенцы, дети и подростки, подвергшиеся воздействию для взрослых из категорий высокого риска
Пятнадцать мм и более положительны в
Ложноположительный результат
Некоторые люди могут реагировать на ТКП, даже если они не инфицированы M.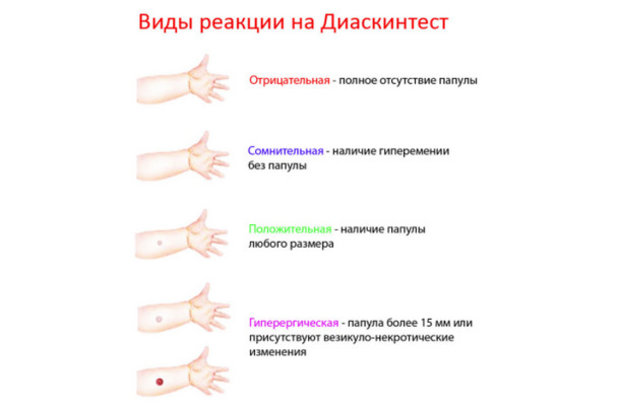 Туберкулез . Причины этих ложноположительных реакций могут включать в себя, помимо прочего, следующее:
Туберкулез . Причины этих ложноположительных реакций могут включать в себя, помимо прочего, следующее:
Инфекция нетуберкулезными микобактериями
Предыдущая вакцинация БЦЖ
Неправильный метод проведения TST
Неправильный интерпретация реакции
Использован неправильный флакон с антигеном
Из-за низкой специфичности теста большинство положительных реакций у лиц с низким риском являются ложноположительными.[8] Ложноположительный результат может быть вызван нетуберкулезными микобактериями или предыдущим введением вакцины БЦЖ. Предыдущая вакцинация БЦЖ может привести к ложноположительному результату в течение многих лет [9].
Ложноотрицательный результат
Отрицательный результат Манту обычно означает, что человек никогда не подвергался воздействию M. tuberculosis . Однако есть факторы, которые могут вызвать ложноотрицательный результат или снижение способности реагировать на туберкулин. [10,11]
Кожная анергия (анергия — это неспособность реагировать на кожные пробы из-за ослабленной иммунной системы)
Недавняя инфекция ТБ (в течение 8-10 недель после заражения)
Очень старая инфекция ТБ (много лет)
Очень молодой возраст (менее шести месяцев)
Недавняя вакцинация живым вирусом (е.ж., корь и оспа)
Одностороннее заболевание туберкулезом
Некоторые вирусные заболевания (например, корь и ветряная оспа)
Неправильный метод проведения ТКП
Неправильная интерпретация реакции, недостаточная доза и непреднамеренная подкожная инъекция.
Отсутствие клеточного иммунитета к туберкулину может быть связано с отсутствием предшествующей сенсибилизации или с ложноотрицательным результатом по разным причинам или с анергией из-за подавления иммунитета.Большинство детей с отрицательным результатом не были инфицированы M. tuberculosis . Небольшая часть в остальном нормальных детей с инфекцией M. tuberculosis остается PPD-отрицательной по неизвестным причинам. С момента заражения до развития CMI существует период окна продолжительностью от двух до шести недель, когда проба Манту будет отрицательной. Пациенты с ослабленным иммунитетом, особенно с ВИЧ и низким числом Т-лимфоцитов CD4, часто показывают отрицательные результаты теста PPD.Это связано с тем, что иммунная система должна быть функциональной, чтобы реагировать на производное белка, вводимое под кожу.
Отрицательные тесты можно интерпретировать как то, что человек не был инфицирован бактериями ТБ или что человек был инфицирован недавно, и прошло недостаточно времени, чтобы организм отреагировал на кожный тест. Повторный тест не рекомендуется ранее, чем через неделю, поскольку туберкулин, вводимый для первого теста, оказывает бустер-эффект на последующую дозу. TST может перейти в положительный результат ≤ восьми недель после заражения Mycobacterium tuberculosis , этот интервал обычно называют «периодом окна». Отрицательный результат ТКП, полученный менее чем за восемь недель до этого, не исключает инфекции, и второй тест рекомендуется через восемь недель. [12] Кроме того, поскольку развитие реакции у пожилого человека может занять более 72 часов, может оказаться полезным повторить кожную пробу на ТБ через 96 часов и еще раз через одну неделю для адекватного скрининга этих людей. Люди с ослабленным иммунитетом могут быть не в состоянии адекватно отреагировать на пробу Манту, и может потребоваться либо рентген грудной клетки, либо образец мокроты.
Отрицательный результат ТКП, полученный менее чем за восемь недель до этого, не исключает инфекции, и второй тест рекомендуется через восемь недель. [12] Кроме того, поскольку развитие реакции у пожилого человека может занять более 72 часов, может оказаться полезным повторить кожную пробу на ТБ через 96 часов и еще раз через одну неделю для адекватного скрининга этих людей. Люди с ослабленным иммунитетом могут быть не в состоянии адекватно отреагировать на пробу Манту, и может потребоваться либо рентген грудной клетки, либо образец мокроты.
Интерпретация для детей: Правильно проведенная проба Манту может иметь неоценимое значение при обследовании ребенка с подозрением на ТБ.Однако интерпретация результата часто затруднена, так как разные работники используют уплотнения разного размера, чтобы указать на положительную реакцию. Хотя сам тест не является ни на 100% ни чувствительным, ни на 100% специфичным, прогностическая ценность положительной реакции очень высока в такой группе. Ранее было показано, что недоедание влияет на результаты туберкулиновой пробы. Как и в других исследованиях, у детей с недостаточным весом в этом исследовании вероятность получения отрицательного результата по Манту была значительно выше [].
Ранее было показано, что недоедание влияет на результаты туберкулиновой пробы. Как и в других исследованиях, у детей с недостаточным весом в этом исследовании вероятность получения отрицательного результата по Манту была значительно выше [].
Таблица 1
Пороговая величина реакции при положительном результате пробы Манту у детей
Как и во многих других исследованиях, у большинства детей не было реакции на туберкулин, несмотря на то, что они получили иммунизацию БЦЖ вскоре после рождения. Причины этого не всегда ясны, но ясно, что любая чувствительность к туберкулину, которую могла вызвать БЦЖ, не могла быть значительной или устойчивой. Это согласуется с нынешней рекомендацией о том, что для пациентов с высоким риском туберкулеза история вакцинации БЦЖ не должна учитываться при интерпретации туберкулиновой пробы.
УСИЛИТЕЛЬНЫЙ ЭФФЕКТ
У некоторых людей, инфицированных M. tuberculosis , способность реагировать на туберкулин со временем может уменьшаться. При проведении ТКП через несколько лет после заражения у этих людей может быть ложноотрицательная реакция. Однако TST может стимулировать иммунную систему, вызывая положительную или повышенную реакцию на последующие тесты. Проведение второй TST после первоначальной отрицательной реакции TST называется двухэтапным тестированием. Когда сенсибилизация к микобактериям произошла много лет назад, первоначальная внутрикожная инъекция туберкулина может вызвать отрицательный или слабоположительный ответ из-за того, что в кровотоке слишком мало сенсибилизированных лимфоцитов, чтобы вызвать значительный местный ответ.Если тест повторяется, большее значение может быть получено из-за того, что иммунный ответ «отзывается» или «усиливается» первым тестом. Второе усиленное показание является правильным, то есть результатом, который следует использовать для принятия решения или будущего сравнения. Повышение является максимальным, если второй тест проводится между одной и пятью неделями после первоначального теста, и он может продолжаться до двух лет.
При проведении ТКП через несколько лет после заражения у этих людей может быть ложноотрицательная реакция. Однако TST может стимулировать иммунную систему, вызывая положительную или повышенную реакцию на последующие тесты. Проведение второй TST после первоначальной отрицательной реакции TST называется двухэтапным тестированием. Когда сенсибилизация к микобактериям произошла много лет назад, первоначальная внутрикожная инъекция туберкулина может вызвать отрицательный или слабоположительный ответ из-за того, что в кровотоке слишком мало сенсибилизированных лимфоцитов, чтобы вызвать значительный местный ответ.Если тест повторяется, большее значение может быть получено из-за того, что иммунный ответ «отзывается» или «усиливается» первым тестом. Второе усиленное показание является правильным, то есть результатом, который следует использовать для принятия решения или будущего сравнения. Повышение является максимальным, если второй тест проводится между одной и пятью неделями после первоначального теста, и он может продолжаться до двух лет.
РЕВЕРСИЯ МАНТУ
Реверсирование определяется как изменение отрицательного результата Манту после предыдущего положительного результата.Как правило, это явление не характерно для здоровых людей и встречается менее чем у 10% таких людей с ранее положительным диагнозом Манту.
Реверсия более распространена [13]
у пожилых людей (по оценкам, 8% в год)
, когда исходное значение Манту <14 мм
у тех, у кого исходная положительная реакция была усилена результат (определяется двухэтапным тестированием).
КОНВЕРСИЯ МАНТУ
В то время как бустинг — это отзыв реакции гиперчувствительности в отсутствие новой инфекции, конверсия — это развитие новой или повышенной гиперчувствительности из-за инфекции туберкулезными или нетуберкулезными микобактериями, включая вакцинацию БЦЖ.
Конверсия Манту определяется [14] как изменение (в течение двухлетнего периода) реактивности Манту, которое соответствует любому из следующих критериев:
изменение с отрицательной реакции на положительную
увеличение ≥ 10 мм.
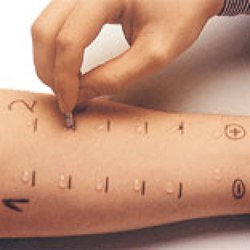
Конверсия была связана с ежегодной заболеваемостью туберкулезом 4% среди подростков [15] или 6% среди лиц, контактировавших с пациентами с положительным мазком [16].
Споры ведутся о времени, необходимом для иммунологических изменений, вызывающих конверсию Манту после инфекции.После непреднамеренной вакцинации M. tuberculosis (катастрофа в Любеке) у детей развились положительные реакции в течение трех-семи недель. Другие исследования показали клиническое заболевание с положительным туберкулиновым тестом в период от 19 до 57 дней после заражения, в среднем 37 дней. [1]
Следовательно, при проверке контактных лиц на туберкулез на предмет конверсии второй туберкулиновый тест проводится через восемь недель после даты последнего контакта с больным-источником. (В прошлом использовался традиционный период окна или интервал в 12 недель.)
ВАКЦИНА КАЛЬМЕТТА-ГЕРИНА И ТЕСТ МАНТУ
Существуют разногласия относительно роли теста Манту у вакцинированных людей. Согласно рекомендациям США, ТКП не противопоказана людям, вакцинированным БЦЖ, и что предшествующая вакцинация БЦЖ не должна влиять на интерпретацию теста.
Согласно рекомендациям США, ТКП не противопоказана людям, вакцинированным БЦЖ, и что предшествующая вакцинация БЦЖ не должна влиять на интерпретацию теста.
В соответствии с руководящими принципами США диагностика и лечение латентной туберкулезной инфекции (ЛТБИ) и лечение ЛТИ рассматриваются для любого вакцинированного БЦЖ человека, чья кожная проба составляет 10 мм или больше, при наличии любого из этих обстоятельств:
Был в контакте с другим человеком, больным инфекционным ТБ
Родился или жил в стране с высокой распространенностью ТБ
Постоянно контактирует с группами населения с высокой распространенностью ТБ.
СИТУАЦИИ, КОТОРЫЕ НЕ РЕКОМЕНДУЕТСЯ ТЕСТИРОВАНИЕ МАНТУ
Тест Манту не рекомендуется в следующих ситуациях:
Прошлые реакции Манту ≥ 15 мм: повторение теста не даст новой диагностической информации и создаст дискомфорт
Перенесенное заболевание туберкулезом: не будет получена полезная диагностическая информация и вероятен значительный дискомфорт
Младенцы до 12 недель: положительная реакция очень важна, но отрицательная реакция может указывать на то, что ребенок слишком мал, чтобы дать ответ , и тест необходимо будет повторить, если облучение произошло.
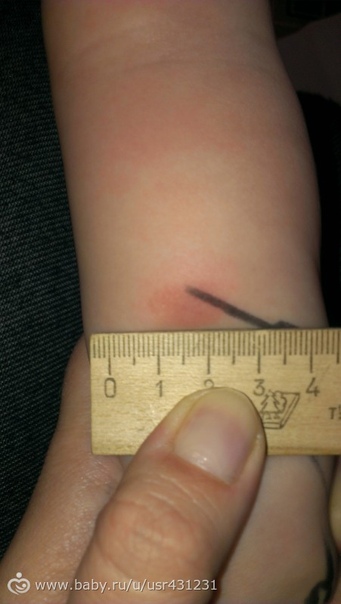 Тестирование Манту перед вакцинацией до достижения 12-недельного возраста не требуется, если ребенок не инфицирован туберкулезом.
Тестирование Манту перед вакцинацией до достижения 12-недельного возраста не требуется, если ребенок не инфицирован туберкулезом.
ПОСЛЕДНИЕ РАЗРАБОТКИ
Тест Манту технически сложен для проведения и чтения, поэтому могут возникнуть ложные показания, если тестировщик не обладает достаточными навыками. Если проводится двухэтапный тест, может потребоваться четыре посещения пациента, а соблюдение этого условия иногда затруднено. Было бы проще сделать анализ, который можно сделать во время одного посещения пациента, например анализ крови.
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов FDA одобрило новый диагностический тест (QuantiFERON-TB GOLD, производимый Cellestis, Inc.) на туберкулез. Анализ крови выявляет наличие инфекции Mycobacterium tuberculosis (TB) путем измерения интерферона-гамма (IFN-G), собранного в плазме цельной крови, инкубированной со специфическими антигенами M. tuberculosis , ESAT-6 (QFT-RD1 ) и CFP-10.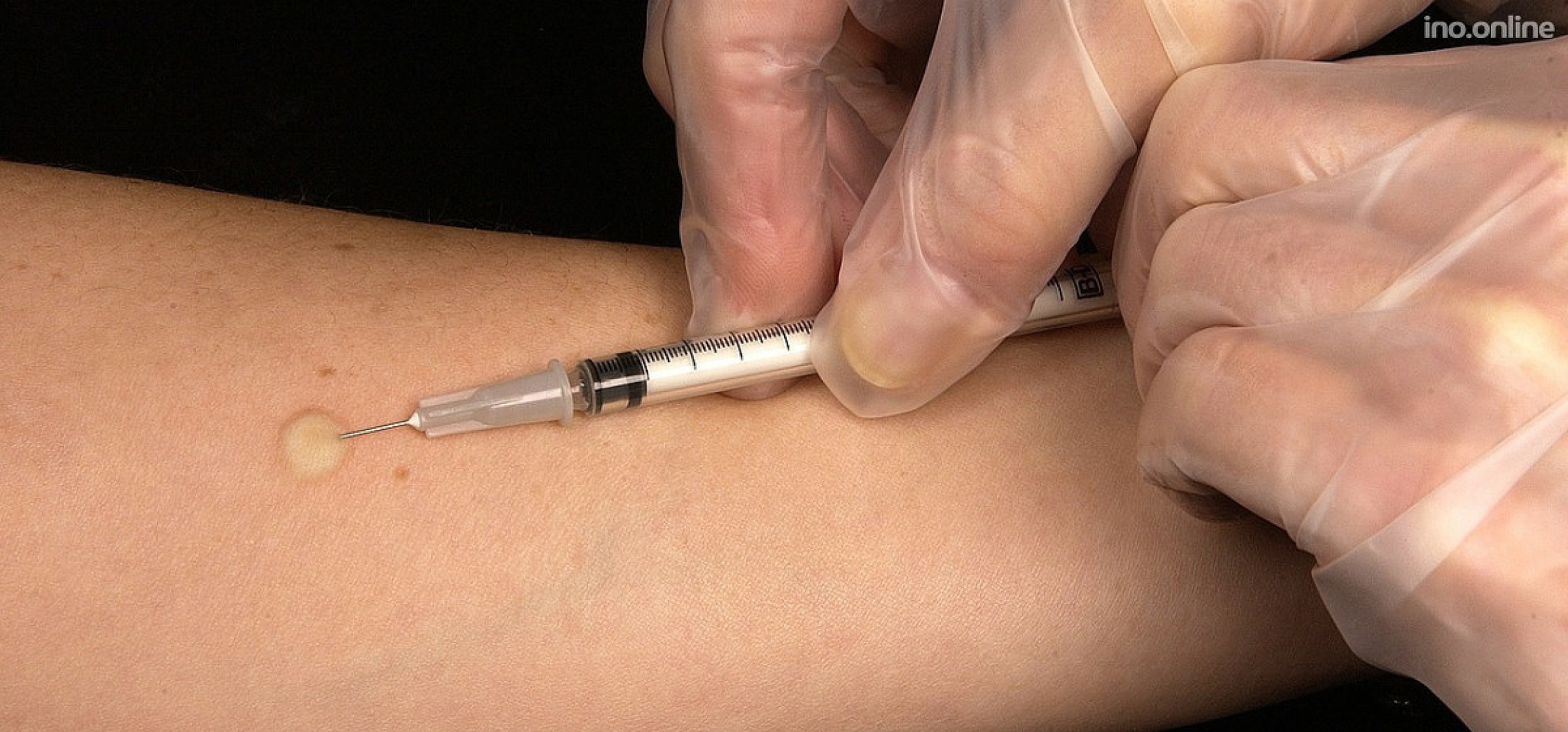 Этот новый иммунодиагностический тест был запущен в качестве вспомогательного средства в диагностике ЛТИ. Комбинация ESAT-6 и CFP10 оказалась высокочувствительной и специфичной как для диагностики in vivo, и in vitro, так и для диагностики .У людей комбинация имела высокую чувствительность (73%) и гораздо более высокую специфичность (93%), чем PPD (7%). Тест QFT-RD1 чувствителен для диагностики ТБ, особенно у пациентов с отрицательными результатами микроскопии и посева. Несмотря на то, что антигены, такие как ESAT-6 и CFP10, не ограничиваются M. tuberculosis , они обещают специфическое обнаружение инфекции M. tuberculosis и могут быть очень полезным дополнительным инструментом для диагностики туберкулеза. .
Этот новый иммунодиагностический тест был запущен в качестве вспомогательного средства в диагностике ЛТИ. Комбинация ESAT-6 и CFP10 оказалась высокочувствительной и специфичной как для диагностики in vivo, и in vitro, так и для диагностики .У людей комбинация имела высокую чувствительность (73%) и гораздо более высокую специфичность (93%), чем PPD (7%). Тест QFT-RD1 чувствителен для диагностики ТБ, особенно у пациентов с отрицательными результатами микроскопии и посева. Несмотря на то, что антигены, такие как ESAT-6 и CFP10, не ограничиваются M. tuberculosis , они обещают специфическое обнаружение инфекции M. tuberculosis и могут быть очень полезным дополнительным инструментом для диагностики туберкулеза. .
Рекомендации по использованию теста QuantiFERON были выпущены [Центром контроля заболеваний] в декабре 2005 года.В настоящее время позиция КвантиФЕРОН-ТБ в диагностике ЛТИ не ясна. Возможно, в будущем появится возможность заменить кожную пробу этим или альтернативным анализом in vitro .
Сноски
Источник поддержки: Нет
Конфликт интересов: Не объявлен.
СПИСОК ЛИТЕРАТУРЫ
1. Мензис Д. Туберкулиновая кожная проба. В: Райхман Л.Б., Хершфилд Е.С., редакторы. Туберкулез: комплексный международный подход. Нью-Йорк: Марсель Деккер; 2000 г.С. 279–322. [Google Scholar] 2. Ховард А., Мерсер П., Натарадж ХК, Канг BC. Скос вниз лучше скоса вверх при внутрикожном кожном тестировании. Ann Allergy Asthma Immunol. 1997. 78: 594–6. [PubMed] [Google Scholar] 3. Американское торакальное общество. Кожная туберкулиновая проба, 1981. Am Rev Respir Dis. 1981; 124: 346–51. [Google Scholar] 4. Риск серьезных аллергических реакций после введения туберсола [очищенного производного белка туберкулина (Манту)] [письмо уважаемому специалисту в области здравоохранения] Оттава: Министерство здравоохранения Канады; 2005 г.Санофи Пастер Лимитед, Отделение продуктов здравоохранения и продуктов здравоохранения Министерства здравоохранения Канады, Агентство общественного здравоохранения Канады. [Google Scholar] 5. Ким Й., Доус-Хиггс Э., Загарелла С. Реакция на инородное тело с участком пробы Манту. Australas J Dermatol. 2005. 46: 169–71. [PubMed] [Google Scholar] 6. Аль Захрани К., Аль Джахдали Х., Мензис Д. Имеет ли значение размер? Полезность величины туберкулиновых реакций для диагностики микобактериальных заболеваний. Am J Resp Crit Care Med. 2000. 162: 1419–22. [PubMed] [Google Scholar] 7. Американское торакальное общество.Заявление о туберкулиновой кожной пробе Американского торакального общества, Медицинской секции Американской ассоциации легких. Am Rev Respir Dis. 1981; 124: 356–63. [Google Scholar] 8. Старке-младший. Кожные пробы на туберкулез: новые школы мысли. J Am Acad Pediatr. 1996; 98: 123–5. [PubMed] [Google Scholar] 9. Чатурведи Н., Кокрофт А. Скрининг на туберкулез среди медицинских работников: кому нужны рентгеновские снимки грудной клетки? Оккуп Мед (Лондон) 1992; 42: 179–82. [PubMed] [Google Scholar] 10. Американское торакальное общество. Стандарты диагностики и классификация туберкулеза у взрослых и детей.Am J Respir Crit Care Med. 2000; 161: 1376–95. [PubMed] [Google Scholar] 11. Американское торакальное общество / Центры контроля заболеваний. Целевое туберкулиновое тестирование и лечение скрытой туберкулезной инфекции. Am J Respir Crit Care Med. 2000; 161: S221–47. [PubMed] [Google Scholar] 12. Anibarro L, Trigo M, Villaverde C, Pena A, Cortizo S, Sande D и др. Анализ высвобождения интерферона-γ у лиц, контактировавших с туберкулезом: есть ли период окна? Eur Respir J. 2011; 37: 215–7. [PubMed] [Google Scholar] 13. Мензис Д. Интерпретация повторных туберкулиновых проб.Повышение, преобразование и возврат. Am J Respir Crit Care Med. 1999; 159: 15–21. [PubMed] [Google Scholar] 14. Целевое туберкулиновое тестирование и лечение скрытой туберкулезной инфекции. Американское торакальное общество. MMWR Recomm Rep. 2000; 49: 1–51. [PubMed] [Google Scholar] 15. Сазерленд И. Эволюция клинического туберкулеза у подростков. Бугорок. 1966; 47: 308. [Google Scholar] 16. Veening GJ. Долгосрочная профилактика изониазидом. Контролируемое испытание по профилактике изониазидом после недавней конверсии туберкулина у молодых людей.Bull Int Union Tuberc. 1968; 41: 169–71. [PubMed] [Google Scholar]Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или уточнить у системного администратора.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файлах cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Аминоацилирование тРНКИ и роль воды в качестве редактора
Abstract
Вычитка / редактирование в синтезе белка имеет важное значение для точного перевода информации из генетического кода. В этой статье мы представляем теоретическое исследование эффективности кинетического механизма корректуры, который использует гидролиз неправильного субстрата в качестве дискриминационного шага в ферментативных каталитических реакциях. Мы рассматриваем аминоацилирование тРНК Ile , которое является важным этапом в синтезе белка и для которого теперь доступны экспериментальные результаты.Мы представляем расширенную кинетическую схему, а затем применяем методы алгоритма стохастического моделирования для получения зависящих от времени концентраций различных веществ, участвующих в реакции, и скорости их образования. Мы получаем скорости образования продукта и гидролиза АТФ как для правильных, так и для неправильных субстратов (изолейцина и валина в нашем случае, соответственно), в одномолекулярном ферменте, а также в кинетике ансамбля ферментов. Настоящая теоретическая схема правильно воспроизводит (i) амплитуду фактора различения в общих скоростях между изолейцином и валином, который получается как (1.8 × 10 2 ). (4,33 × 10 2 ) = 7,8 × 10 4 , (ii) скорости гидролиза АТФ как для Ile, так и для Val при различных концентрациях субстрата при аминоацилировании тРНК Ile . Настоящее исследование показывает зависимость скорости реакции немихайлового типа от концентрации тРНК Ile в случае валина. Установлено, что общее редактирование в устойчивом состоянии не зависит от концентрации аминокислот. Интересно отметить, что рассчитанная скорость гидролиза АТФ для валина при высокой концентрации субстрата такая же, как и скорость образования Ile-тРНК Ile , тогда как при промежуточной концентрации субстрата скорость гидролиза АТФ относительно низкая.Мы обнаружили, что наличие дополнительного редактирующего домена в редактирующем ферменте класса I делает кинетическую корректуру более эффективной за счет усиленного гидролиза неправильного продукта в редактирующем CP1-домене.
Образец цитирования: Сантра М., Багчи Б. (2013) Кинетическая корректура на одномолекулярном уровне: аминоацилирование тРНК Ile и роль воды как редактора. PLoS ONE 8 (6): e66112. https://doi.org/10.1371/journal.pone.0066112
Редактор: Анил Кумар Тьяги, Университет Дели, Индия
Поступила: 16 марта 2013 г .; Одобрена: 2 мая 2013 г .; Опубликован: 20 июня 2013 г.
Авторское право: © 2013 Santra, Bagchi.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Работа частично поддержана грантами DST и CSIR (Индия). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что конкурирующих интересов не существует.
Введение
Кинетическая корректура — это теория, предложенная для объяснения известного отсутствия ошибок в биологическом синтезе. В биохимических реакциях ферменты не только увеличивают скорость реакции, но и выборочно выбирают правильный субстрат, ведущий к желаемому продукту. Многие биологические процессы, такие как синтез белка или репликация ДНК, проявляют высокую специфичность в отношении выбора правильных субстратов в присутствии многих других структурно или химически аналогичных субстратов [1].Ожидается, что из-за сходной энергии связывания как правильных, так и неправильных субстратов и аналога размера / формы фермента частота ошибок (отношение скорости образования неправильного продукта к скорости образования желаемого продукта) будет высокой. Напротив, частота ошибок при выборе аминокислоты при синтезе белка (10 −4 ) [2] и репликации ДНК (10 −9 ) [3] — [5] чрезвычайно низка. Молекулярная причина такой высокой селективности до сих пор не совсем понятна с точки зрения количественной теории.Эта важная проблема оставалась предметом дискуссий в течение нескольких десятилетий, при этом первоначальная формулировка повторной активации Хопфилда была признана неадекватной в нескольких биосинтезах [6]. Недавние экспериментальные исследования нескольких ферментативных каталитических реакций показали, что разложение промежуточных продуктов происходит посредством реакции гидролиза [7], [8]. Было предложено несколько альтернативных механизмов редактирования, которые были признаны удовлетворительными в разных случаях, подчеркивая тот факт, что может работать более одного механизма [9].Один из этих механизмов, впервые предложенный Ферштом, использует гидролиз не того субстрата в качестве основного дискриминационного шага.
В предыдущем исследовании мы исследовали каталитический процесс аминоацилирования тРНК. Мы показали, что для синтетаз класса I стационарные и одиночные молекулярные события дают несколько разную кинетику [10]. Мы представили расширенную кинетическую схему, которая включала роль воды и использовала универсальную технику распределения времени первого прохода для получения зависящей от времени скорости.Теория объясняет несоответствие между однократным оборотом и скоростью устойчивого состояния для ферментов как класса I, так и класса II [10].
В данной работе мы изучаем кинетическую корректуру фермента класса I. Мы изучили относительные достоинства двух схем, предложенных Хопфилдом и Ферштом, и выяснили скорость. Чтобы количественно оценить кинетическую корректуру, мы исследовали с доступными данными степень правильного и неправильного формирования продукта.
В своем новаторском исследовании Фершт предположил, что наиболее важным отличительным шагом в кинетической корректуре может быть гидролиз высокоэнергетического комплекса фермент-субстрат.Таким образом, решающую роль играет простая химическая реакция с водой. Однако подробный количественный анализ схемы Фершта, насколько нам известно, не проводился.
Здесь мы представляем такой анализ. Мы предлагаем схему, подобную Michealis-Menten, в которую мы включаем гидролиз как побочную реакцию, чтобы показать, что гидролиз может помочь ферменту различать два аналогичных субстрата, несмотря на то, что они имеют схожие энергии связывания.
Элементарные стадии ферментативного катализа включают образование комплекса Михаэлиса-Ментен с последующим образованием продукта.В последние годы несколько экспериментальных исследований ферментативного катализа на одномолекулярном уровне выявили много интересных особенностей в краткосрочной динамике [11]. Он успешно воспроизводит ту же скорость стационарного состояния, которую дает кинетика Михаэлиса-Ментен. При одиночном молекулярном ферментативном катализе в системе присутствует только один фермент, и постоянный поток субстрата поддерживается, поскольку концентрация субстрата относительно высока по сравнению с концентрацией фермента. В экспериментальных установках in vitro концентрация фермента остается постоянной за счет рециркуляции свободных ферментов после высвобождения продукта, тогда как концентрация субстрата со временем уменьшается.Ситуация in vivo может быть иной, если концентрация фермента может быть аналогична концентрации субстрата. В настоящей работе мы рассматриваем как реакции одиночного молекулярного фермента, так и реакции ансамблевого ферментного катализа.
В этой статье мы в основном концентрируемся на аминоацилировании тРНК Ile с использованием изолейцина в качестве правильного и валина в качестве неправильного субстрата в присутствии изолейцил-тРНК синтетазы (IleRS) в процессе синтеза белка. Эта система была тщательно изучена в нескольких экспериментах [9], [12], и есть много интересных открытий (в основном кинетических результатов), которые еще предстоит решить.Например, относительная скорость аминоацилирования валина и изолейцина сильно зависит от температуры. Проверка на ∼10 4 не очевидна из-за разницы в скорости гидролиза только продуктов.
С другой стороны, хотя в литературе существует ограниченное количество теоретических исследований, они в большинстве своем не могут объяснить существующие экспериментальные результаты без какого-либо противоречия. В последние годы в связи с развитием передовых методов молекулярной спектроскопии были предприняты огромные попытки проанализировать структуры различных промежуточных продуктов, образующихся в ходе реакции и фермента [13], [14].Хотя структура различных промежуточных продуктов, образующихся в ходе реакции, может раскрыть механизм, она не может дать нам количественную оценку дискриминации. Более того, часто бывает крайне сложно получить все промежуточные продукты, поскольку многие из них временные. Кинетические экспериментальные исследования имеют преимущество количественной оценки скорости отдельной стадии, а также общей реакции. Следовательно, механизм реакции можно понять, построив схему реакции на основе кинетических экспериментальных данных, которая может объяснить все доступные кинетические результаты.Лучший способ понять реакцию как количественно, так и качественно — это предложить схему, основанную как на кинетических, так и на спектральных результатах, и подтвердить экспериментальные наблюдения с результатами, полученными из схемы.
Структурный анализ аминоацил-тРНК-синтетаз показывает, что существует активный сайт, который активирует аминокислоту в присутствии АТФ, а затем активированная аминокислота переносится на тРНК. Уникальность изолейцил-тРНК синтетазы (IleRS) заключается в том, что кроме этого активного сайта у нее есть отдельный сайт редактирования, расположенный на ∼30 Å отдельно от сайта активации в домене, известном как домен CP1 ( рис.1 ) [13], [15] — [17]. Роль сайта редактирования заключается в гидролизе неправильного комплекса фермент-продукт, что приводит к снижению частоты ошибок. Присутствие этого сайта редактирования заставляет IleRS различать валин более эффективно, чем это наблюдается у нередактирующих ферментов аминоацилирования тРНК [18].
Контроль скорости гидролиза АТФ важен, поскольку он напрямую связан со скоростью гидролиза промежуточных продуктов, образующихся во время реакции. Более высокая скорость гидролиза АТФ указывает на более высокую скорость гидролиза промежуточных продуктов, которая служит побочной реакцией, приводящей к образованию меньшего количества продукта.Если гидролиз промежуточных продуктов не происходит, то количество гидролизованных молекул АТФ на молекулу образовавшегося продукта будет идентично 1. Величина отношения, наблюдаемая в случае изолейцина, составляет ~ 1,5, тогда как в случае валина ~ 270 [12]. Таким образом, гидролиз АТФ служит маркером для оценки точности предложенной кинетической схемы.
Сравнение апо Thermus thermophilus LeuRS с сокристаллической структурой комплекса аминоацилирования Pyrococcus horikoshii LeuRS с тРНК-Leu показывает, что домен CP1 поворачивается на ~ 20 ° относительно канонического ядра фермента, чтобы предотвратить конфликт со связанным 5 ‘Конец тРНК [15].После аминоацилирования 3’-конец тРНК перемещается на ~ 30 Å от синтетического активного сайта к сайту редактирования аминокислоты для проверки; требует, чтобы область редактирования повернулась на ~ 35 ° по сравнению с апо-состоянием T. thermophilus LeuRS.
Накопление многочисленных кристаллических структур LeuRS ясно показывает, что фермент претерпевает значительные конформационные изменения, чтобы приспособиться к комплексам аминоацилирования и редактирования. Это также сопровождается изменениями в тРНК, особенно на заряженном 3′-конце, и ее взаимодействиями с AARS.
Сокристалл S. aureus IleRS с тРНК Ile показал, что сайт редактирования внутри CP1 может быть доступен с 3′-конца связанной тРНК Ile . Было предложено конформационное изменение связанной тРНК для перемещения 3′-конца от активного сайта к сайту редактирования.
Элегантный анализ кинетической корректуры, объясняющий повышенную специфичность цикла фосфорилирования-дефосфорилирования (PdPC), был недавно проведен Qian [19]. PdPC демонстрирует сверхчувствительность нулевого порядка и, как известно, демонстрирует усиление чувствительности.PdPC также может распознавать неспецифические перекрестные помехи при передаче сигнала. Фактически, многие кинетические механизмы корректуры зависят от механизма обратной связи, который приводит к переключению, описывающему усиление чувствительности.
Предлагаемые кинетические схемы и их описание
В альтернативной схеме Фершта [9] повторная активация не требуется. Здесь критическим шагом является повышение скорости гидролиза неправильного субстрата. Фершт описал два механизма, как показано на рис.2 и Рис.3 .
Рис. 2. Схематическое изображение одного из двух кинетических механизмов корректуры (механизм I), предложенных Ферштом.
Здесь образование Val-тРНК Ile является лимитирующей стадией (k = 1,2 с -1 ), за которой следует быстрый гидролиз комплекса.
https://doi.org/10.1371/journal.pone.0066112.g002
Рис. 3. Альтернативный механизм кинетической корректуры (механизм II), предложенный Ферштом.
В этом механизме изменение конформации фермента является самым медленным шагом. Общая кинетика включает гидролиз в два этапа.
https://doi.org/10.1371/journal.pone.0066112.g003
Согласно механизму I ( рис. 2 ), в случае валина фермент переходит в сильно гидролизуемое состояние во время образования IleRS *. .Val-AMP.tRNA Иль комплекс. После этапа определения скорости , т.е. ., переноса валина на тРНК Ile комплекс быстро гидролизуется, приводя к свободному ферменту и исходным субстратам.Таким образом, предполагается, что гидролиз увеличивает дискриминацию с 10 до ∼180. В альтернативном механизме, показанном на фиг.3 (механизм II), Фершт предположил, что существует возможность фермента претерпевать конформационные изменения и переходить в высоко гидролизуемую конформацию еще до переноса валина на тРНК Ile (образуя IleRS ). * .Val-AMP.tRNA Ile ), и это этап ограничения скорости. После конформационной флуктуации комплекс Val-AMP.tRNA Ile быстро гидролизуется.Существует еще одна стадия гидролиза, связанная с продуктом (Val-тРНК Ile ), если он образован по ошибке. На основании результатов экспериментов Фершт пришел к выводу, что оба этих механизма равновероятны.
Однако с обоими механизмами есть некоторые сложности. В обоих случаях предполагается, что фермент претерпевает изменение конформации до высокогидролизуемого состояния. Структурный анализ IleRS показывает, что он имеет сайт редактирования, расположенный в домене CP1, который находится на ∼30 Å от активного сайта (см. Рис.1 ), а валин перемещается в этот сайт после переноса на тРНК Ile и легко гидролизуется [13], [20] — [26]. Это говорит о том, что фермент не должен подвергаться каким-либо конформационным колебаниям. Скорее, внутри фермента есть сайт связывания, и субстрат переходит к этому сайту и гидролизуется. Поскольку эта транслокация происходит после образования связи Val-тРНК Ile , стадия гидролиза участвует после образования Val-тРНК Ile , как в механизме I.Это возражает против механизма II.
Теперь, согласно механизму I, фермент переходит в гидролизуемое состояние во время образования комплекса Val-AMP.IleRS * .tRNA Ile и гидролизуется после образования связи Val-тРНК Ile . Этап определения скорости считается этапом перед гидролизом , т.е. ., этапом, который включает образование связи между аминокислотой и тРНК. Однако несколько экспериментальных свидетельств подтверждают, что стадия выпуска продукта является стадией ограничения скорости.
Таким образом, ни один из вышеперечисленных механизмов не обеспечивает должного гидролиза. Здесь мы конструируем механизм, основанный как на кинетических экспериментах, так и на структурных доказательствах, и проводим количественное исследование, чтобы понять вклад усиленного гидролиза в общую дискриминацию и гидролиз АТФ. Отметим, что такой количественный анализ не проводился даже для механизмов Фершта.
На основе экспериментальных результатов Фершта и Хопфилда мы построили новую теоретическую модель, которая имеет следующие особенности.
- Гидролиз происходит только после образования продукта, связанного с ферментом, но до высвобождения продукта.
- Этап образования фермент-субстратного комплекса (ES) включает небольшую дискриминацию (~ 1/10) [27], как в схеме Хоупфилда. Однако реакция этого комплекса с АТФ включает дальнейшее несоответствие, приводящее к общему различению 1/160 до образования комплекса фермент-субстрат-АМФ.
- В случае неправильного переноса субстрата (валина) на тРНК Ile , аминокислота попадает в домен CP1 (гидролитически активная область) фермента.В результате он подвергается быстрой диссоциации за счет усиленного гидролиза.
- Чрезвычайно низкая концентрация комплекса фермент-продукт в устойчивом состоянии, обнаруженная Ферштом в случае валина, предполагает, что он подвергается усиленному гидролизу.
После учета всех вышеперечисленных наблюдений, мы разработали новую схему, которая показана на Рис. 4 (для изолейцина).
Предлагаемая модифицированная схема реакции валина немного отличается и изображена на рис.5 .
На рис. 4 и рис. 5 и . , IleRS обозначено как E , это аминокислота, это АТФ, это АМФ, это тРНК Ile , это продукт (или или), указывает, что продукт находится в домене CP1. Мы использовали две немного разные схемы для Ile и Val. Введение дополнительных стадий гидролиза в случае Val связано с экспериментальными доказательствами гидролиза Val на соответствующих стадиях. Обратите внимание, что основное различие между этими двумя схемами заключается в наличии транслокации Val-тРНК Ile в редактирующий домен CP1, приводящей к образованию высокогидролизуемого комплекса E * P в случае Val.
Теперь мы определяем зависящую от времени скорость катализа и концентрации гидролиза АТФ для обоих субстратов. Вышеупомянутые кинетические схемы могут быть представлены системой простых химических кинетических уравнений. Решение этих уравнений обеспечивает зависящую от времени концентрацию субстратов, различных промежуточных продуктов и продуктов, из которых можно легко определить скорость катализа и гидролиза АТФ. Поскольку представленные схемы сложны, аналитическое решение получить слишком сложно.Хотя можно найти решение для заданного набора кинетических параметров с помощью метода численного интегрирования, решение, полученное этим методом, является детерминированным по своей природе. С другой стороны, реакции in vivo не являются детерминированными из-за небольшого размера биологической клетки. Существует альтернативный метод получения недетерминированного решения, следующий за методом распределения времени первого прохождения. Этот метод следует вероятностному подходу, который находит распределение времени первого прохождения образования продукта, из которого легко может быть получена скорость установившегося режима.Два основных ограничения этого подхода следующие. Для сложных реакций, включающих различные элементарные стадии, такие как настоящие кинетические схемы, это не работает. Другой и наиболее важный недостаток заключается в том, что он не обеспечивает зависимость концентраций субстратов, промежуточных соединений и продуктов от времени, поскольку он имеет дело с вероятностью времени первого прохождения, а не с вероятностями веществ.
Методы
Мы провели стохастический симуляционный анализ, предложенный Гиллеспи, и получили зависящие от времени концентрации всех субстратов, промежуточных соединений и продуктов [28].Это недетерминированный метод, который позволяет определять концентрации различных веществ с течением времени. В то время как для небольшой системы он показывает случайную концентрацию во времени, для большой системы получается детерминированная зависящая от времени концентрация. Более того, с помощью этого метода были изучены как одиночный молекулярный, так и ансамблевый ферментный катализ.
Обсуждение и результаты
Приведенные выше кинетические схемы переводятся в систему кинетических уравнений, знакомую из химической кинетики, особенно в области кинетики ферментов.Существует несколько методов решения такой связанной системы дифференциальных уравнений первого порядка. Один из подходов, который мы использовали ранее, — это метод распределения времени первого прохождения (f (t)) путем непосредственного вывода уравнения для f (t). Этот метод практичен, когда количество связанных уравнений невелико, например, шесть или меньше. Однако здесь мы имеем дело с большим количеством концентраций, более десяти. Итак, мы использовали метод стохастического моделирования Гиллеспи для получения зависимых от времени концентраций [28].Этот метод и его различные варианты широко обсуждались в литературе.
Для проведения анализа необходимы числовые значения всех кинетических параметров. Мы рассмотрели следующий набор k ’s для изолейцина, которые либо получены, либо взяты из литературы [9], [12], [27].
,,,,, ,,, и.
Аналогично, константы скорости для валина равны
.,,,,,,,, ,,, и.
Во-первых, мы исследовали одиночный молекулярный катализ для обоих субстратов.В этом исследовании рассматривается один фермент, а другие субстраты (аминокислоты, АТФ и тРНК Ile ) берутся в избыточном количестве, что аналогично методике экспериментов с одной молекулой. Рис. 6 и Рис. 7 показывают распределения времени ожидания для Ile и Val соответственно. Рисунки ясно показывают, что временные масштабы образования продукта для изолюцина и валина сильно различаются. Скорость образования продукта идентична распределению времени первого прохождения.Распределение времени первого прохождения показывает, что скорость образования продукта изолейцина очень быстро снижается и стремится к нулю в течение секунды. С другой стороны, соответствующая скорость для валина не сильно меняется даже до 10 секунд (показано на вставке , рис. 7, ). Экспериментальное исследование также показывает подобное поведение [12]. Большая разница во времени для двух субстратов возникает из-за присутствия гидролиза перед переносом и после переноса в случае валина.
Рисунок 7.Вычисленное распределение времени первого прохождения для аминоацилирования тРНК Ile валином в одномолекулярной реакции.
На вставке показана временная зависимость на коротком промежутке времени, где распределение показывает сначала резкий подъем, а затем медленный спад.
https://doi.org/10.1371/journal.pone.0066112.g007
Роль тРНК Ile в гидролизе АТФ для получения валина представлена на рис. 8 . Это показывает, что скорость гидролиза АТФ в стационарном состоянии увеличивается с увеличением концентрации тРНК Ile .При низкой концентрации тРНК Ile она быстро увеличивается, а затем насыщается при высокой концентрации. Аналогичное поведение наблюдается в случае изолейцина (данные не показаны). Стационарная скорость гидролиза АТФ как для валина (, фиг. 9 (A), ), так и для изолейцина (, фиг. 9 (B), ) показывает поведение типа Михаэлиса-Ментен. Интересно, что график обратной скорости по сравнению с обратной концентрацией тРНК Ile в случае валина (вставка , рис.9 (A), ) указывает на отклонение от поведения Михаэлиса-Ментен в отличие от изолейцина (вставка рис.9 (В) ). Это отклонение объясняется корреляцией относительного вклада тРНК-независимого гидролиза в тРНК-зависимый гидролиз. Таким образом, наличие тРНК-независимого гидролиза усложняет зависимость общего гидролиза АТФ от тРНК Ile .
Рис. 9. Зависимость от концентрации тРНК Ile стационарной скорости гидролиза АТФ для (A) валина и (B) изолейцина.
На вставках показано соответствующее поведение в обратных масштабах.Обратите внимание на отклонение от поведения Михаэлиса-Ментен для изолейцина.
https://doi.org/10.1371/journal.pone.0066112.g009
Большинство недавних экспериментальных исследований по кинетической корректуре аминоацилирования тРНК пытаются понять относительный вклад редактирования до и после переноса, а также тРНК-независимое и зависимое редактирование, так как механизм редактирования отражается на этих величинах. Незначительный вклад тРНК-независимого редактирования посредством гидролиза валина наблюдался неоднократно, на что указывает скорость гидролиза АТФ в отсутствие тРНК Ile .Однако экспериментально невозможно оценить относительный вклад тРНК-независимого редактирования в присутствии тРНК Ile . В нашем настоящем исследовании мы пометили молекулы AMP, которые образуются в результате гидролиза на стадии редактирования, независимой от тРНК. Как тРНК-зависимая, так и независимая скорость гидролиза АТФ представлена в Fig. 10 . Как и ожидалось, скорость тРНК-зависимого гидролиза АТФ увеличивается с увеличением концентрации тРНК Ile . Интересно, что скорость тРНК-независимого гидролиза АТФ снижается с увеличением концентрации тРНК Ile и спадает до нуля при высокой концентрации.Следует отметить, что хотя тРНК-независимый гидролиз АТФ приближается к нулю, относительный вклад редактирования перед переносом уменьшается, а затем сходится к конечному значению (~ 9%) при высокой концентрации тРНК Ile .
Рисунок 10. ТРНК Ile , зависящая от концентрации стационарная скорость гидролиза АТФ для Val в процессах до и после переноса.
Доля гидролиза перед переносом по отношению к общему гидролизу АТФ также показана синей пунктирной линией.Относительный вклад редактирования перед переносом сходится к конечному значению (∼9%) при высокой концентрации тРНК Ile .
https://doi.org/10.1371/journal.pone.0066112.g010
Уменьшение относительного вклада тРНК-независимого редактирования происходит из-за того, что популяция фермент-тРНК Ile увеличивается, а свободный фермент уменьшается с увеличение концентрации тРНК Ile . При очень высокой концентрации тРНК Ile почти все ферменты остаются в виде комплекса фермент-тРНК Ile , тогда как тРНК-независимое редактирование происходит только тогда, когда фермент начинает каталитический цикл в свободной форме.На фиг. , фиг. 11, мы показываем фракцию свободного фермента и комплекса фермент-тРНК Ile при двух предельных концентрациях тРНК Ile . При низкой концентрации ферменты остаются в обеих формах. С другой стороны, при высокой концентрации он показывает значительное количество комплекса фермент-тРНК Ile по сравнению со свободным ферментом.
Рисунок 11. Зависимость от времени доли свободного фермента и комплекса фермент-тРНК Ile при двух различных концентрациях тРНК Ile ([тРНК Ile ] = 1 нМ и 100 нМ) в условиях многооборотного обращения в присутствии валина. .
Пунктирная и сплошная линии соответствуют концентрациям тРНК Ile 1 нМ и 100 нМ соответственно. Черные и зеленые линии указывают на долю свободного фермента, тогда как красные и синие линии указывают на долю комплекса фермент-тРНК.
https://doi.org/10.1371/journal.pone.0066112.g011
Стационарные скорости катализа изолейцина и валина и гидролиза АТФ для валина показаны на Рис. 12 . Зависимость скорости реакции от концентрации аминокислот для обоих субстратов демонстрирует поведение, подобное Михаэлису-Ментену.Хотя скорость гидролиза АТФ для валина отличается от скорости реакции для изолейцина при малых и промежуточных концентрациях аминокислот, они почти равны при высоких концентрациях аминокислот и насыщаются до значения ∼1,1 с -1 .
Рис. 12. Зависимость от концентрации аминокислот стационарной скорости ферментативного катализа для изолейцина (черная сплошная линия) и валина (синяя пунктирная линия).
Скорость гидролиза АТФ для Val показана красной пунктирной линией.
https://doi.org/10.1371/journal.pone.0066112.g012
Было обнаружено, что скорость аминоацилирования изолейцина выше, чем скорость гидролиза АТФ в случае валина в промежуточной концентрации (72 мкМ) субстрат [12]. При очень высокой концентрации субстрата другое экспериментальное исследование показывает, что эти две скорости абсолютно одинаковы при различных температурах и pH системы [9]. Важно отметить, что хотя скорость аминоацилирования изолейцина такая же, как и скорость гидролиза АТФ в случае валина при высокой концентрации субстрата, первая скорость всегда выше, чем вторая при концентрации аминокислот от низкой до промежуточной.Из зависимости скорости образования продукта в установившемся состоянии для изолейцина и валина от концентрации аминокислот мы находим, что константы Михаэлиса составляют ∼60 нМ и ∼26 мкМ соответственно. В результате в диапазоне микромолярных концентраций установившаяся скорость образования продукта изолейцина становится насыщенной, но гидролиз АТФ для валина все еще остается ниже своего максимального значения. Разница в шкале констант Михаэлиса для изолейцина и валина снижает скорость гидролиза АТФ для валина по сравнению со скоростью образования продукта изолейцина в промежуточном диапазоне концентраций (до 500 мкМ).Скорость образования продукта валина остается низкой во всем диапазоне концентраций аминокислот (, фиг. 12, ).
Здесь необходимо упомянуть, что стационарная концентрация Val-тРНК Ile составляет менее 0,8% от комплекса IleRS или IleRS.Val-AMP. [9] Как высокое значение отношения АТФ к продукту, так и низкая концентрация Val-тРНК Ile предполагают, что скорость гидролиза во время реакции намного выше, чем нормальная скорость гидролиза Val-тРНК Ile .Это возможно, если аминокислота попадает в сильно гидролизующий домен CP1 IleRS после переноса на тРНК Ile . Включение такого усиленного гидролиза во время реакции объясняет как высокое значение соотношения, так и низкую концентрацию Val-тРНК Ile .
Общее различие, когда оба субстрата присутствуют в реакционной смеси в равном количестве, представляет собой отношение k cat / K m для неправильных и правильных субстратов. Таким образом, дискриминация.В настоящем исследовании мы получили ( k cat ) Ile = 1.057 s −1 и ( k cat ) Val = 5,76 × 10 −3 s −1 . Таким образом, соотношение ( k cat ) Ile / ( k cat ) Val равно 1,8 × 10 2 . Мы также получили значения ( K м ) Ile и ( K м ) Val , и это 6.17 × 10 −8 M и 2,68 × 10 −5 M соответственно. Тогда отношение ( K м ) Ile / ( K м ) Val получается равным 4,33 × 10 2 . Здесь следует отметить, что экспериментально определенное значение ( K м ) Ile / ( K м ) Val составляет ∼1 / 10 2 [27]. Следовательно, общая дискриминация составляет (1,8 × 10 2 ).(4,3 × 10 2 ) = 7,8 × 10 4 .
До сих пор мы представили кинетику изолейцина и валина и соответствующее редактирование, рассматривая субстраты (изолейцин и валин) отдельно. Чтобы получить полное редактирование напрямую, мы проводим анализ в присутствии обоих субстратов с равным количеством в условиях насыщения тРНК , Ile и АТФ. Зависящие от времени продукты высеваются на левой панели Рис. 13 . Рисунок и соответствующая вставка показывают, что общее редактирование составляет ∼10 5 , что близко к вышеупомянутому значению D.Мы обнаружили, что редактирование в установившемся состоянии не зависит от концентрации аминокислот, присутствующих в системе (правая панель , рис. 13, ).
Рисунок 13. Левая панель: зависимость продуктов от времени (Ile-тРНК Ile и Val-тРНК Ile ), когда и изолейцин, и валин присутствуют в системе в равных количествах.
На вставке показано линейное увеличение Val-тРНК Ile во время устойчивого состояния. Правая панель: зависимость от концентрации аминокислот общего редактирования ( i.е. , соотношение скоростей образования Ile-тРНК Ile и Val-тРНК Ile в присутствии равных количеств Ile и Val).
https://doi.org/10.1371/journal.pone.0066112.g013
Заключение
Учитывая множество процессов, участвующих в аминоацилировании тРНК, количественное понимание кинетической корректуры непросто. Наличие множества констант скорости также затрудняет точное экспериментальное объяснение механизма корректуры.Таким образом, цель состоит не только в том, чтобы различать схемы Хопфилда и Фершта, но и в рационализации констант скорости, наблюдаемых экспериментально.
Достижения в области спектроскопии одиночных молекул вселяют надежду, поскольку можно получить распределение времени ожидания, которое содержит больше информации, чем просто скорость стационарного состояния. Также существует возможность, что мы можем наблюдать отдельные процессы отдельно и индивидуально.
В этой статье мы представили модифицированную схему Фершта, чтобы количественно понять происхождение эффективного кинетического считывания при аминоацилировании тРНК Ile , рассматривая конкретный случай различения изолейцина и валина.Схема Фершта предлагает гидролиз как ключевой шаг различения. С помощью предложенной нами схемы мы получили распределение времени первого прохождения, а также стационарную скорость реакции и гидролиза АТФ для Ile и Val. Настоящее теоретическое исследование успешно фиксирует экспериментальные наблюдения, так что разница в скорости общей реакции для Ile и Val. Мы показываем, что стационарная скорость гидролиза АТФ при высокой концентрации Val такая же, как и скорость образования продукта для Ile при его высокой концентрации, тогда как при низкой и промежуточной концентрации аминокислот скорость гидролиза АТФ для валина меньше, чем скорость Ile Образование -тРНК Ile .Общая скорость реакции в стационарном состоянии в случае валина показывает зависимость типа, отличного от michaelis, от концентрации тРНК Ile из-за взаимодействия между тРНК-независимым и тРНК-зависимым редактированием. Вклад тРНК-независимого редактирования в общее редактирование валина уменьшается с увеличением концентрации тРНК Ile и стремится к нулю при высокой концентрации. Мы обнаружили, что значительное количество фермента остается в виде комплекса фермент-тРНК Ile по сравнению со свободным ферментом, и это снижает относительный вклад независимого от тРНК редактирования.Настоящая работа дает количественную оценку важности этапа гидролиза и обнаруживает, что в разумной кинетической схеме этот этап может в общих чертах объяснить величину дискриминации при кинетической корректуре.
Есть еще много важных этапов, связанных с синтезом белка, таких как кинетическая корректура, участвующая в отборе аминоацил-тРНК в рибосоме [29], [30] и выборе правильной пары оснований для репликации ДНК. Трудность формулировки правильной кинетической схемы для любой из этих задач заключается в отсутствии констант скорости для нескольких критических шагов.Таким образом, даже когда предложена правильная схема, ее справедливость не может быть легко подтверждена. Требуются дальнейшие экспериментальные исследования отдельных кинетических ступеней.
Молекулярный механизм редактирования еще предстоит понять на микроскопическом уровне. Ясно, что каталитические домены CP1 и Rossmann fold могут определять ход событий, следующих за образованием продукта. Возможно, что размер не того продукта позволяет ему перейти в домен CP1, в то время как правильный продукт будет выпущен без перехода в домен CP1.Недавний структурный анализ выявил доказательства конформационного движения домена CP1 с большой амплитудой вокруг глицина на β-цепи. Одномолекулярное спектроскопическое исследование, безусловно, может помочь в выяснении правильного механизма.
Благодарности
Мы хотели бы поблагодарить профессора Умеша Варшнея за обсуждения и за указание ценных ссылок. Мы благодарим профессора Алана Фершта за полезные предложения. М.С. благодарит г-на Ракеша Шаран Сингха, Раджиба Бисваса и Сайката Банерджи за ценный вклад в подготовку рукописи.
Вклад авторов
Задумал и спроектировал эксперименты: MS BB. Проведены эксперименты: ср. Проанализированы данные: MS BB. Написал статью: MS BB.
Список литературы
- 1. Полинг Л. (1958) В: Festschrift Arthur Stoll, Birkhäuser Verlag, Базель, Швейцария, стр.597.
- 2. Ленингер А.Л. (1970) Биохимия: молекулярные основы структуры и функции клетки. Нью-Йорк: Worth Publishers, 833 с.
- 3. Уотсон Дж. Д. (1970) Молекулярная биология гена.Нью-Йорк: В. А. Бенджамин, 662 с.
- 4. Кимура М., Охта Т. (1973) Мутация и эволюция на молекулярном уровне. Genetics Suppl 73: 19–35.
- 5. Фершт А.Р. (1981) Обзорная лекция: Механизмы редактирования ферментов и генетический код. Proc R Soc Lond B 212: 351–379.
- 6. Хопфилд Дж. Дж. (1974) Кинетическая корректура: новый механизм уменьшения ошибок в биосинтетических процессах, требующий высокой специфичности. Proc Natl Acad Sci U S A 71: 4135–4139.
- 7. Хуссейн Т., Камартапу В., Крупарани С.П., Дешмук М.В., Санкаранараянан Р. (2010) Механистические взгляды на распознавание родственных субстратов во время корректуры при переводе. Proc Natl Acad Sci U S A 107: 22117–22121.
- 8. Boniecki MT, Vu MT, Betha AK, Martinis SA (2008) CP1-зависимое разделение предварительного и посттрансферного редактирования в лейцил-тРНК-синтетазе. Proc Natl Acad Sci U S A 105: 19223–19228.
- 9. Фершт А.Р. (1977) Механизмы редактирования в синтезе белков.Отторжение валина изолейцил-тРНК синтетазой. Биохимия 16: 1025–1030.
- 10. Santra M, Bagchi B (2012) Катализ аминоацилирования тРНК: одиночный оборот к стационарной кинетике тРНК синтетаз. J. Phys Chem B 116: 11809–11817.
- 11. Лу HP, Xun L, Xie XS (1998) Одномолекулярная ферментативная динамика. Science 282: 1877–1882.
- 12. Hopfield JJ, Yamane T, Yue V, Coutts SM (1976) Прямое экспериментальное доказательство кинетической корректуры при аминоацилировании тРНК Ile .Proc Natl Acad Sci U S A 73: 1164–1168.
- 13. Фукаи С., Нуреки О, Секин С., Шимада А., Тао Дж. И др. (2000) Структурная основа для разделения L-валина на L-изолейцин и L-треонин с помощью двойного сита с помощью комплекса тРНК val и валил-тРНК синтетазы. Cell 103: 793–803.
- 14. Mascarenhas AP, Martinis SA (2008) Функциональная сегрегация предсказанного «шарнирного» сайта в линкерах β-цепи лейцил-тРНК синтетазы Escherichia coli .Биохимия 47: 4808–4816.
- 15. Zhu B, Yao P, Tan M, Eriani G, Wang E (2009) tRNA-независимое предварительное редактирование с помощью лейцил-тРНК синтетазы класса I. J Biol Chem 284: 3418–3424.
- 16. Silvian LF, Wang J, Steitz TA (1999) Взгляд на редактирование структуры Ile-тРНК-синтетазы с тРНК Ile и мупироцином. Science 285: 1074–1077.
- 17. Кьюсак С., Яремчук А., Тукало М. (2000) Кристаллическая структура 2 Å лейцил-тРНК синтетазы и ее комплекса с аналогом лейциладенилата.EMBO J 19: 2351–2361.
- 18. Gruic-Sovulj I, Uter N, Bullock T, Perona JJ (2005) тРНК-зависимый аминоацил-аденилатный гидролиз нередактирующей аминоацил-тРНК синтетазой класса I. J Biol Chem 280: 23978–23986.
- 19. Qian H (2003) Термодинамический и кинетический анализ усиления чувствительности при передаче биологического сигнала. Biophy Chem 105: 585–593.
- 20. Splan KE, Игнатов ME, Musier-Forsyth K (2008) Трансферная РНК модулирует механизм редактирования, используемый пролил-тРНК синтетазой класса II.J Biol Chem 283: 7128-7134.
- 21. Апостол I, Левин Дж., Липпинкотт Дж., Лич Дж., Хесс Э. и др. (1997) Включение норвалина в положения лейцина в рекомбинантном человеческом гемоглобине, экспрессируемом в Escherichia coli . J Biol Chem 272: 28980–28988.
- 22. Чжао М., Чжу Б., Хао Р., Сюй М., Эриани Г. и др. (2005) Лейцил-тРНК-синтетаза из предковой бактерии Aquifex aeolicus содержит реликты эволюции синтетазы. EMBO J 24: 1430–1439.
- 23. Фершт А.Р. (1985) Структура и механизм фермента. Нью-Йорк: В. Х. Фриман, 475 с.
- 24. Eriani G, Delarue M, Poch O, Gangloff J, Moras D (1990) Разделение тРНК-синтетаз на два класса на основе взаимоисключающих наборов мотивов последовательностей. Природа 347: 203–206.
- 25. Номанбхой Т.К., Хендриксон Т.Л., Шиммель П. (1999) Зависимая от переносимой РНК транслокация неправильно активированных аминокислот для предотвращения ошибок в синтезе белка.Mol Cell 4: 519–528.
- 26. Nomanbhoy TK, Schimmel PR (2000) Мизактивированные аминокислоты перемещаются с одинаковой скоростью по поверхности тРНК-синтетазы. Proc Natl Acad Sci U S A 97: 5119–5122.
- 27. Лофтфилд Р. Б., Эйгнер Е. А. (1966) Специфика ферментативных реакций. Аминоацилрастворимые РНК-лигазы. Biochim Biophys Acta 130: 426–448.
- 28. Gillespie DT (1976) Общий метод численного моделирования стохастической временной эволюции связанных химических реакций.Журнал Comput Phys 22: 403–434.
- 29. Blanchard SC, Gonzalez RL, Kim HD, Chu S, Puglisi JD (2004) Выбор тРНК и кинетическая корректура при переводе. Nat Struct Mol Biol 11: 1008–1014.
- 30. Schmeing TM, Ramakrishnan V (2009) Что недавние структуры рибосом показали о механизме трансляции. Природа 461: 1234–1242.
| 190 | Семинар 2018 21 декабря 2018 — 12:00 Нобелевская премия по физике 2018 Проф. Дебабрата Госвами, Индийский технологический институт Канпур |
| 189 | Lecture2018 07 декабря 2018 — 16:00 Открытая лекция на тему «План жизни» Проф.Бенни Шило, Институт Вейцмана, Израиль |
| 188 | Seminar2018 05 декабря 2018 — 23:00 Основная матрица мономиальной кривой и что она может рассказать нам о кривой Проф. Хема Сринивасан |
| 187 | Семинар 2018 3 декабря 2018 г. — 16:00 Локальная унификация, дефекты и связанные с ними ранжированные кольца Проф. Дейл Каткоски (Университет Миссури) |
| 186 | Seminar2018 29 ноя 2018 — 17:00 Электронная структура новых двумерных материалов Sanjoy K.Mahatha, Deutsches Elektronen-Synchrotron DESY, Гамбург, Германия |
| 185 | Seminar2018 28 ноября 2018 — 16:00 Стратегическое регулирование фотосинтетического механизма на пути к генерированию биоэнергии и биопереработке для максимального извлечения продукта Доктор Рашми Чандра, профессор-исследователь, Tecnologico de Monterrey, Школа инженерии и наук, Campus Toluca Tecnologico de Монтеррей, Мексика |
| 184 | Seminar2018 27 ноября 2018 г. — 17:00 Металлоорганические соединения: понимание, применение и возможности Д-р Видья Авасаре, факультет химии, Колледж сэра Парашурамбхау, Пуна |
| 183 | Seminar2018 27 ноя 2018 — 15:30 Дистальные действия на сферах Dr.Алок Кумар (J.N.U.) |
| 182 | Seminar2018 27 ноября 2018 — 15:00 Наследие в современной Индии: исследование сикхских музеев Каника Сингх (доктор философии, Университет Амбедкара, Дели) |
| 181 | Seminar2018 26 ноября 2018 — 17:00 Пластичность и механическая текучесть в аморфных твердых телах Прабхат Джайсвал, Институт инженерии и технологий Thapar, Пенджаб |
| 180 | Seminar2018 26 ноября 2018 — 16:00 Конституция как инструмент реализации идеи Индии: исторический анализ законодательных и судебных вмешательств (1946-2018) V.Кришна Анант (профессор исторического факультета Университета SRM, Амаравати) |
| 179 | Семинар 2018 26 ноября 2018 — 15:00 Изучить молекулярный механизм метаболизма некодирующей РНК для симметричной мутации в локусе IGH Д-р Панкадж Кумар Гири, Колумбийский университет, Нью-Йорк (США) |
| 178 | Семинар 2018 26 ноября 2018 г. — 10:30 Угроза мобильности, загробная жизнь наследственной преступности и межпоколенческой проституции в сообществе Субир Рана (доктор философии, Университет Джавахарлала Неру, Дели) |
| 177 | Seminar2018 22 ноя 2018 — 17:00 Стратегии селективной C─H-функционализации для синтеза бензоконденсированных гетероциклов Проф.Перунчералатан, Школа химических наук, Национальный институт научного образования и исследований (NISER) Бхубанешвар |
| 176 | Семинар 2018 22 ноября 2018 — 15:00 Ближайшие галактики и их окружение Смрити Махаджан |
| 175 | Seminar2018 22 ноября 2018 — 15:00 Об исчезающих телах и воплощенных знаках: телесность в цифровой фазе Раджни Муджрал (доктор философии, Университет английского и иностранных языков, Хайдарабад) |
| 174 | Seminar2018 22 ноября 2018 — 12:00 Специальный семинар по эмиссии микробных летучих органических соединений бактериями и грибами Dr.Павел Мишталь, группа ученых 6), в настоящее время работает в Центре экологии и гидрологии НКРЭ, Эдинбург, | .
| 173 | Seminar2018 19 ноября 2018 — 15:00 Оценка измерений аэрозолей черного углерода в Арктическом регионе Доктор Пунья Р. Синха Научный сотрудник, D, Balloon Facility @ Tata Institute of Fundamental Research, Хайдарабад |
| 172 | Seminar2018 16 ноября 2018 — 18:00 Органические превращения с использованием ацеталей, образованных in situ Проф. Ренгараджан Баламуруган, Школа химии Хайдарабадского университета |
| 171 | Seminar2018 16 ноя 2018 — 16:00 Архитектура и сила: понимание экономической эффективности с 2014 Проф.Пулапре Балакришнан, Университет Ашока, Сонипат |
| 170 | Семинар 2018 16 ноября 2018 — 15:00 Микробы — устойчивый источник биоматериалов, биоэнергии и биоактивных молекул Доктор Шаши Кант Университет Бхатиа Конкук, Сеул, Южная Корея |
| 169 | Seminar2018 16 ноября 2018 г. — 10:30 От целлулоида к цифровым: мир жизни бходжпури СМИ Акшая Кумар (доктор философии, Университет Глазго) |
| 168 | Семинар 2018 15 ноя 2018 — 17:00 Детекторная аппаратура для исследований в области ядерной физики Давиндер Сивал, факультет физики, Университет Пенджаба |
| 167 | Seminar2018 14 ноя 2018 — 17:00 Асимптотические симметрии в гравитации и плоской голографии Рудранил Басу, Université Libre de Bruxelles |
| 166 | Seminar2018 13 ноября 2018 — 17:00 Квантовая материя в редкоземельных пирохлорах и полуклассический подход к квантовому ограничению хаоса Самбудда Саньял, Колумбийский университет, Нью-Йорк |
| 165 | Seminar2018 09 ноя 2018 — 15:00 Роль SNARE-опосредованного экзоцитоза и аутофагии в патогенезе панкреатита Dr.Субханкар Долай, Университет Торонто, |
| 164 | Seminar2018 05 ноя 2018 — 15:00 Излечить безумие? Социальная и культурная история безумия в колониальной Индии, 1800-1947 годы Шилпи Раджпал (доктор философии, Университет Дели) |
| 163 | Seminar2018 01 ноя 2018 — 17:00 Нобелевская премия по медицине 2018 г .: личная точка зрения! Проф. Дипанкар Нанди, Департамент биохимии, IISc Bangalore |
| 162 | Семинар 2018 30 октября 2018 — 15:00 Промышленность, связи и информатизация: чайные плантации в рамках глобальной производственной системы в Индии Дебдулал Саха (PhD, TISS Mumbai) |
| 161 | Seminar2018 29 октября 2018 г. — 15:00 Экологизм маргинализированных: постколониальное экологическое прочтение избранных рассказов о глобальном Юге Нибедита Бандйопадхьяй (PhD, IIT Kanpur) |
| 160 | Seminar2018 26 окт 2018 — 17:00 Единый подход к стохастическим сравнениям многомерных моделей смесей Dr.Самин Накви (Китайский университет Гонконга) |
| 159 | Семинар 2018 26 октября 2018 — 15:00 Рассказы о взаимодействии паразита и хозяина со стороны самого смертоносного из врагов человека — комара Доктор Маюр Кумар Каджла Рамалингасвами Сотрудник / Независимый ИП ICMR-Национальный институт исследований малярии, Нью-Дели, Индия . |
| 158 | Seminar2018 24 октября 2018 — 17:00 Новые подходы к характеристике динамики, электростатики, структуры и сворачивания белков Dr.Рамкришна Адхикари, Исследовательский институт Скриппса, Калифорния, США |
| 157 | Семинар2018 24 октября 2018 г. — 16:00 Количественная оценка запасов подземных вод и разработка биогеохимической модели Д-р Сумендра Нат Бханджа, научный сотрудник, факультет науки и технологий, Университет Атабаски, Эдмонтон, Канада |
| 156 | Seminar2018 24 октября 2018 — 15:00 Новое байесовское определение подтверждения и парадокс неуместного соединения Филоза Коши (доктор философии, Хайдарабадский университет) |
| 155 | Seminar2018 12 окт 2018 — 17:00 О кривых положительного решения бесконечной полупозитонной задачи Dr.Дханья Раджендран (ИИТ Гоа). |
| 154 | Seminar2018 12 октября 2018 — 15:00 Метаболизм в разных шкалах и его перекрестная связь с клеточными сигналами Д-р Виней Булусу, Онкологические исследования UK Beatson Institute, Глазго, Великобритания |
| 153 | Seminar2018 08 октября 2018 — 15:00 О здоровье и здравоохранении: путешествие антрополога и эволюция исследовательских идей Ретиндер Каур (доктор философии, Пенджабский университет) |
| 152 | Seminar2018 05 окт 2018 — 17:00 О гиперболических законах сохранения и законах сохранения с прерывным потоком Dr.Сударшан Кумар К. |
| 151 | Seminar2018 05 октября 2018 — 15:00 Фантастические микробы и их местонахождение: когда и как ученые заимствуют данные из литературы Нираджа Шанкаран |
| 150 | Семинар 2018 05 октября 2018 — 15:00 Двойная роль Swi6 (гетерохроматиновый белок 1) в контроле мейотической рекомбинации и сегрегации вокруг центромер Д-р Мридула Намбиар, Отдел фундаментальных наук, Центр исследования рака Фреда Хатчинсона, Сиэтл, США |
| 149 | Seminar2018 04 окт 2018 — 17:00 Сверхчувствительное обнаружение наноматериалов без этикеток и терапевтическое применение Dr.Сударсон Сехар Синха, Школа физики и астрономии, Тель-Авивский университет, Израиль |
| 148 | Семинар 2018 04 окт 2018 — 17:00 Квантовое моделирование с ультрахолодными атомами в оптических решетках Бодхадитья Сантра |
| 147 | Семинар 2018 4 октября 2018 г. — 15:00 Противотуберкулезные кампании в постколониальной Южной Индии и Юго-Восточной Азии: пути, не избранные в международном здравоохранении, 1950-е годы Вивек Нилкантан (доктор философии, Сиднейский университет) |
| 146 | Seminar2018 02 октября 2018 — 16:00 Полуэмпирический подход к моделированию сильного движения грунта: перспектива оценки сейсмической опасности Dr.Сандип Доцент кафедры геофизики Института науки БХУ Варанаси |
| 145 | Семинар 2018 2 октября 2018 — 15:00 The Beat Revolt Ашима Бхардвадж (доктор философии, Университет Пенджаба) |
| 144 | Seminar2018 01 октября 2018 — 15:00 Демографический переход, «избыток» мужчин и проблема межпоколенческого контракта в Северной Индии Паро Мишра (доктор философии, ИИТ Дели) |
| 143 | Conference / Workshop2018 01 октября 2018 — 08:00 GIAN Курс по структурированному взаимодействию света и материи и его применению Проф.Этьен Брасселе, Университет Бордо, Франция, |
| 142 | Seminar2018 28 сен 2018 — 17:00 Создание последовательностей и полугрупп оценок Д-р Арпан Датта |
| 141 | Семинар 2018 28 сен 2018 — 17:00 Лакказы и другие: новые катализируемые медью превращения в органическом синтезе Проф. Уве Бейфус, Университет Хоэнхайм, Штутгарт, Германия |
| 140 | Лекция 2018 27 сентября 2018 — 16:00 Foundation Day Lecture 2018 |
| 139 | Лекция 2018 27 сентября 2018 г. — 09:00 Программа Foundation Day 2018 |
| 138 | Seminar2018 26 сен 2018 — 17:00 Неадиабатическая динамика внутримолекулярного синглетного деления в ковалентно связанных димерах пентацена Dr.Раджагопала Р. Силам, Институт теоретической физики, Университет Эрлангена, Германия |
| 137 | Seminar2018 26 сентября 2018 — 16:00 Вредное цветение водорослей для зеленых технологий в мире, испытывающем антропогенные и климатические изменения Д-р Анкита Шривастава, Исследовательский центр клеточных фабрик, Корейский научно-исследовательский институт биологии и биотехнологии (KRIBB), Тэджон 34141, Республика Корея |
| 136 | Seminar2018 25 сентября 2018 — 16:00 Как понять жизнь и философию Ганди: некоторые предложения Dr.Нишикант Колге, Центр изучения развивающихся обществ (CSDS), Дели. |
| 135 | Семинар 2018 20 сен 2018 — 17:00 Бифункциональность в металлоорганическом катализе Проф. Джитендра К. Бера, Индийский технологический институт Канпур |
| 134 | Seminar2018 19 сен 2018 — 17:00 Разработка катализаторов для клещей на основе сотрудничества металл-лиганд: элегантный подход к устойчивому катализу Dr.Субрата Чакраборти, Институт науки Вейцмана, Реховот, Израиль |
| 133 | Семинар 2018 19 сентября 2018 — 16:00 Рассказы о взаимодействии паразита и хозяина самого смертоносного из врагов человека — комара Доктор Маюр Кумар Каджла, научный сотрудник Рамалингасвами, ICMR-Национальный институт исследований малярии (NIMR) ), Нью-Дели |
| 132 | Seminar2018 18 сен 2018 — 17:00 Химические подходы к ингибированию и активации клинически важных ферментов Проф.Иш Кумар, Школа естественных наук, Университет Фэрли Дикинсон, Нью-Джерси, США |
| 131 | Seminar2018 14 сентября 2018 — 17:00 Спектроскопия и динамика в сверхбыстрой временной области Проф. Дипак К. Палит, Центр передового опыта в области фундаментальных наук UM-DAE, Университет Мумбаи, кампус Калина, Мумбаи |
| 130 | Seminar2018 13 сен 2018 — 17:00 Разработка супрамолекулярных биогибридных и интеллектуальных наноматериалов для применения в нано-био интерфейсах Dr.Sabyasachi Chakrabortty, Институт исследования полимеров Макса Планка, Германия |
| 129 | Seminar2018 11 сентября 2018 — 17:00 Квантовая обработка информации и гиперполяризация ядерных спинов с использованием центров азотных вакансий в алмазе К. Рама Котесвара Рао, Technische Universitaet Dortmund, Дортмунд, Германия |
| 128 | Seminar2018 07 сен 2018 — 17:00 Создание последовательностей и полугрупп оценок Dr.Арпан Дутта |
| 127 | Семинар 2018 06 сен 2018 — 17:00 Активные коллоиды в нематических жидких кристаллах Сатьянараяна Паладугу |
| 126 | Семинар 2018 05 сен 2018 — 17:00 Передовые технологии для открытия материалов, инноваций и их применения Д-р Шриманта Пахира, Индийский технологический институт (ИИТ), Индор |
| 125 | Конференция / семинар2018 01 сентября 2018 — 09:30 Семинар на тему «Подход, ориентированный на решение для сохранения водных ресурсов» |
| 124 | Seminar2018 31 августа 2018 — 17:00 Космическое микроволновое фоновое излучение как инструмент для исследования Вселенной Проф.Т. Р. Сешадри, Делийский университет |
| 123 | Seminar2018 29 августа 2018 — 17:00 Молекулярное понимание механизма липидной модуляции GPCR и мембранной организации / динамики в передаче сигналов Д-р Мутуси Манна, Отдел химии, IISER Bhopal |
| 122 | Семинар 2018 29 августа 2018 г. — 10:00 Катализируемые фосфорной кислотой энантиоселективные превращения Н-связанных орто-хинон-метидов и орто-хинон-метид-иминов Проф Кристоф Шнайдер, Лейпцигский университет, Германия |
| 121 | Seminar2018 28 августа 2018 — 17:00 Пространство Тейхмюллера многомерных многообразий с отрицательной кривизной Dr.Ганготрий Соркар, Университет штата Огайо, |
| 120 | Seminar2018 28 августа 2018 — 17:00 Приближение типа Коровкина в алгебрах C * Д-р Виджайраджан А. К., Школа математики Керла, Кожикоде |
| 119 | Seminar2018 27 августа 2018 — 17:00 Рентгеновские наблюдения: исследование областей с экстремальной гравитацией в рентгеновских двойных системах Ару Бери, Саутгемптонский университет |
| 118 | Seminar2018 24 августа 2018 — 15:30 Соединения с координационными связями C → N Проф.Прасад В. Бхаратам, декан, НИПЕР Мохали |
| 117 | Семинар 2018 24 августа 2018 — 15:00 РНК G-Quadruplex: повторный поиск Проф. Сувик Маити, CSIR-Институт геномики и интегративной биологии, Дели, Индия |
| 116 | Seminar2018 24 августа 2018 г. — 04:30 Специализированные молекулы и сборки для прогнозирования ответов Проф. Амитава Дас, директор CSIR-CSMCRI, Бхавнагар |
| 115 | Seminar2018 23 августа 2018 — 17:00 Моделирование ферментных реакций методами QM / MM Dr.Субхенду Рой, Университет Южной Калифорнии, США |
| 114 | Семинар 2018 23 августа 2018 — 17:00 Биофизика активных мембран с биологическими наномашинами Трипта Бхатия, Институт коллоидов и интерфейсов Макса Планка |
| 113 | Seminar2018 20 августа 2018 — 17:00 О некоторых эллиптических краевых задачах с линейными и квазилинейными операторами Абхишек Саркар (Университет Западной Богемии, Чешская Республика) |
| 112 | Seminar2018 16 августа 2018 — 17:00 Синтезы противомалярийных и противораковых агентов Dr.Рохан Дилипрао Эранде, Университет Райса, Техас, США |
| 111 | Seminar2018 10 августа 2018 — 15:00 Функции и глобальные зависимости факторов сплайсинга делящихся дрожжей предполагают разнообразие в сборке сплайсосом и выборе места сплайсинга Профессор Уша Виджайрагхаван, Отдел микробиологии и клеточной биологии Индийского института науки (IISc) Бангалор |
| 110 | Seminar2018 08 августа 2018 — 17:00 Метастабильное разделение жидкой и жидкой фаз и критичность в воде Dr. p-OE для класса гиперболических групп коксетера Dr.Каджал Дас (Институт Вейцмана, Израиль). |
| 106 | Seminar2018 19 июн 2018 — 16:00 Специальный семинар по топологическому сверхпроводнику и майорановским фермионам Д-р Сурин Дас (IISER Kolkata) |
| 105 | Семинар 2018 06 июня 2018 — 15:00 Делящиеся дрожжи: модельная система для понимания динамики гетерохроматина и коммерческого применения Доктор Джагмохан Сингх, бывший старший научный сотрудник Института микробных технологий, Чандигарх |
| 104 | Seminar2018 01 июн 2018 — 15:00 Искусство разрыва и создания пептидных связей Проф.Р. П. Рой, Национальный институт иммунологии, Нью-Дели |
| 103 | Семинар 2018 31 мая 2018 г. — 11:00 Конфайнмент и скученность контролируют размер, форму и положение бактериальной хромосомы Д-р Дебасиш Чаудхури (IOP, Бхубанешвар) |
| 102 | Seminar2018 28 мая 2018 г. — 16:00 Настройка фотофизики красных флуоресцентных белков Д-р Премашис Манна, JILA, Департамент химии и биохимии, Университет Колорадо, Боулдер |
| 101 | Seminar2018 25 мая 2018 — 15:00 Онкогенная роль локализованной цитоплазмы ARID1B Dr.Мурали Дхаран Башьям, штатный научный сотрудник и руководитель лаборатории молекулярной онкологии, Центр дактилоскопии и диагностики ДНК (CDFD), Хайдарабад |
| 100 | Семинар 2018 23 мая 2018 — 15:00 Патогенное оружие для подрыва защиты хозяина Проф. Амитабха Мукхопадхьяй, НИИ, Нью-Дели |
| 99 | Seminar2018 17 мая 2018 — 16:00 Эквивариантные отображения, связанные с топологической гипотезой Тверберга Dr.Суроджит Гош (ISI Калькутта) |
| 98 | Seminar2018 16 мая 2018 — 11:45 Множественные особенности равновесной свободной энергии в одномерной модели мягких стержней Профессор Дипак Дхар (IISER Pune) |
| 97 | Семинар 2018 16 мая 2018 г. — 10:30 Вырожденные метрики в гравитации первого порядка Профессор Рамеш Каул (IMSc) |
| 96 | Seminar2018 10 мая 2018 — 16:00 Динамика локальных напряжений в суспензиях, утолщающих сдвиг Dr.Викрам Рати (Джорджтаунский университет, Вашингтон) |
| 95 | Seminar2018 09 мая 2018 — 16:00 Структурные фазовые переходы между фазами вращателя нормальных алканов Д-р Прабир К. Мукерджи (Правительственный колледж инженерии и текстильных технологий, Хугли) |
| 94 | Семинар 2018 4 мая 2018 г. — 16:00 Кристаллизация и ее применение на коллекторах PL Д-р Биплаб Басак (Корнельский университет) |
| 93 | Seminar2018 4 мая 2018 — 16:00 Последние исследования столкновений тяжелых ионов при промежуточных энергиях Dr.Сунил Кумар (Университет Чандигарха, Мохали) |
| 92 | Seminar2018 03 мая 2018 — 16:00 Новое доказательство гипотезы Хельгасона первого ранга и приложения к теории рассеяния Д-р Апрамян Партасарати (Университет Падерборна, Германия) |
| 91 | Seminar2018 03 мая 2018 — 16:00 Конденсаты Бозе-Эйнштейна в оптических решетках с искусственными магнитными полями Д-р Раши Сачдева (OIST, Окинава, Япония) |
| 90 | Seminar2018 02 мая 2018 — 16:00 Визуализация магнитных структур: от источников микропотоков до управляемой вихревой материи Dr.Горки Шоу (Льежский университет, Бельгия) |
| 89 | Семинар 2018 01 мая 2018 — 11:00 Слоистые сегнетоэлектрические материалы для устройств памяти Д-р Харихара Венкатараман (BITS-PILANI, Хайдарабад) |
| 88 | Seminar2018 27 апр 2018 — 17:00 Гипотеза Шалева для типа A_n Проф. Анупам Сингх (IISER Pune) |
| 87 | Seminar2018 27 апреля 2018 г. — 11:00 Свойства и приложения нового бета-обобщенного распределения Проф.Канчан Джайн (Департамент статистики, Пенджабский университет) |
| 86 | Семинар 2018 20 апр 2018 — 17:00 Скоро уточняется Д-р Анупам Пал Чоудхури (Институт Иоганна Радона, Австрия) |
| 85 | Семинар 2018 20 апр 2018 — 16:00 История эволюции и происхождение пакетов BIF-основных-ультраосновных-анортозитов в зоне сдвига Мойяр-Бхавани-Кавери (MBCSZ) из гранулитового террейна на юге Индии (GTSI) Dr.Мумика Талукдар, Школа нефтяных технологий, Нефтяной университет Пандит Дендаял, Гандинагар |
| 84 | Семинар 2018 20 апреля 2018 г. — 15:00 Грунтовка для защиты растений: устойчивый подход к защите растений Д-р Прашант Сингх, факультет ботаники, Университет Раджастана |
| 83 | Семинар 2018 20 апр 2018 — 05:00 Нематические жидкие кристаллы в липшицевых областях Д-р Анупам Пал Чоудхури (Институт Иоганна Радона, Австрия) |
| 82 | Seminar2018 19 апр 2018 — 17:00 Гармонические карты и дикие пространства Тейхмюллера Субходжой Гупта (IISC Bangalore) |
| 81 | Seminar2018 19 апр 2018 — 16:00 Петрология пелитовых пород с низким и высоким содержанием барровского типа метаморфизма вокруг Мангпу (Дарджилинг), Гималаев и Кандры (Джаркханд) Dr.Супарна Тевари, Центр наук о Земле, Индийский институт науки, Бангалор, |
| 80 | Семинар 2018 19 апр 2018 — 15:30 Управление динамикой для разработки сложных химических систем Доктор Субхабрата Маити, Университет штата Пенсильвания, США |
| 79 | Семинар 2018 18 апр 2018 — 17:00 Сопоставление по предпочтениям: многомерный подход Д-р Сушмита Гупта (Университет Бергена, Норвегия) |
| 78 | Семинар 2018 18 апр 2018 — 12:00 Проективные структуры на поверхностях Субходжой Гупта (IISC Bangalore) |
| 77 | Seminar2018 17 апр 2018 — 17:00 Раскрытие основных процессов нуклеиновых кислот: роль крио-ЭМ Dr.Индраджит Лахири, Калифорнийский университет, Сан-Диего, |
| 76 | Seminar2018 17 апр 2018 — 17:00 На пути к следующему поколению приложений магнитного резонанса с помощью устойчивой ядерной спиновой гиперполяризации Соумья С. Рой, Йоркский университет, Великобритания |
| 75 | Seminar2018 16 апр 2018 — 17:00 Динамика пассивного полимера на активной подложке Ниша Гупта (IISER Mohali) |
| 74 | Seminar2018 14 апр 2018 — 16:00 Смектики Де Фриза с большим электроклиническим эффектом в фазе SmA * и необычной небольшой усадкой в фазе SmC * — Перспективные материалы для следующего поколения дисплеев FLC Проф.Джагдиш К. Видж, факультет электронной и электротехники, Тринити-колледж, Дублин, Дублинский университет, Ирландия |
| 73 | Конференция / семинар2018 14 апр 2018 — 09:00 Семинар на тему «Передача науки с использованием индийского народного искусства» .. Семинар по народным наукам доктора Битасты Даса и г-на Пратика Прабхакара |
| 72 | Seminar2018 13 апр 2018 — 17:00 Механическое исследование ферментов с потенциалом в терапии и биотопливе Dr.Дебасис Дас, Департамент биологии и Департамент химии, Массачусетский технологический институт, Кембридж, США |
| 71 | Семинар 2018 13 апр 2018 — 17:00 Вырожденные модели Уиттакера и охватывающие группы Д-р Шив Пракаш Патель (IISC Bangalore) |
| 70 | Seminar2018 13 апр 2018 — 16:00 Специальный семинар по роли остеопонтина, хемокина в микросреде опухоли груди: новая парадигма в терапии рака Dr.Гопал Кунду, научный сотрудник и профессор, Национальный центр клеточных исследований, Пуна, |
| 69 | Seminar2018 12 апр 2018 — 16:00 Многопараметрическое моделирование распространения сейсмических поверхностных волн и характеристики b-значения в Северо-восточной Индии Д-р Сантану Манна, Дисциплина гражданского строительства и математика, ИИТ Индор |
| 68 | Seminar2018 11 апр 2018 — 17:00 Чистота для семей репрезентаций Галуа Джиоти Пракаш Саха (Университет Бен-Гуриона, Израиль) |
| 67 | Seminar2018 09 апр 2018 — 17:00 Нелинейный гравитационный коллапс в моделях с темной энергией Проф.Джасджит Сингх Багла (IISER Mohali) |
| 66 | Seminar2018 09 апр 2018 — 15:00 CRISPR-Cas9, управляемая малыми молекулами, в направлении геномной инженерии в клетках Доктор Басудеб Маджи, Институт Броуда Гарварда и Массачусетского технологического института, США |
| 65 | Семинар 2018 06 апр 2018 — 17:00 Понимание квантовых спиновых жидких фаз в фрустрированных магнитах Самбудда Саньял, Колумбийский университет |
| 64 | Seminar2018 6 апр 2018 — 17:00 Арифметические пробелы Dr.Арнаб Саха (ANU Австралия) |
| 63 | Seminar2018 06 апр 2018 — 16:00 Индийский летний муссон в рамках регионального моделирования и ассимиляции данных Д-р Раджу Аттада, Отдел компьютерных, электрических и математических наук и инженерии (CEMSE), Университет науки и технологий имени короля Абдаллы (КАУСТ) ) |
| 62 | Seminar2018 06 апр 2018 — 15:30 Вмешательство в интерференцию: нацеливание на путь РНКи с использованием малых молекул Проф.Сувик Маити, CSIR-Институт геномики и интегративной биологии, Дели |
| 61 | Seminar2018 06 апр 2018 — 15:00 Матрицы, многочлены и повторения над конечными полями Профессор Судхир Р. Горпейд (ИИТ Бомбей) |
| 60 | Seminar2018 05 апр 2018 — 17:00 Спектроскопическое исследование природных явлений: от атмосферных процессов до образования клатратов Д-р Анамика Мухопадхьяй, доцент кафедры физики, Департамент наук, Женский колледж Гуру Гобинд Сингха, Чандигарх |
| 59 | Seminar2018 04 апр 2018 — 17:00 Одноионные магниты типа Изинга как строительные блоки для создания высокоэффективных одномолекулярных магнитов Dr.Арун Кумар Бар, Школа химии, Университет Манчестера, Великобритания |
| 58 | Seminar2018 03 апр 2018 — 17:00 О кольце когомологий грассманиана ориентированных трех плоскостей Д-р Пратип Чакраборти (IISER Kolkata) |
| 57 | Семинар 2018 3 апреля 2018 — 15:00 Регулирование ультраструктуры компонентов ранних секреторных путей Д-р Дибьенду Бхаттачарья, Лаборатория биологии органелл, Advanced Center for Treatment Research & Education in Cancer (ACTREC), Mumbai |
| 56 | Seminar2018 02 апр 2018 — 17:00 Прямые и косвенные исследования новой физики на LHC Dr.Диша Бхатия (TIFR, Мумбаи) |
| 55 | Семинар 2018 27 марта 2018 — 17:00 Структурные аспекты ферритиновой белковой клетки для конструирования неорганических биоматериалов Доктор Басудев Мэйти, Школа естественных наук и технологий, Токийский технологический институт, Япония |
| 54 | Seminar2018 23 марта 2018 — 16:00 Адаптивные вейвлет-методы для решения PDE и анализа сигналов Доктор Ратиканта Бехера (IISER Kolkata) |
| 53 | Seminar2018 23 марта 2018 — 15:00 Эпитранскриптом РНК в Положении о переводе Dr.Намит Ранджан, Отдел физической биохимии, Институт биофизической химии Макса Планка, Геттинген, Германия |
| 52 | Seminar2018 22 марта 2018 — 17:00 Неорганическая химия и катализ: от промышленного применения к операндо-спектроскопии Д-р Абхишек Дутта Чоудхури, научный сотрудник группы неорганической химии и катализа, Утрехтский университет, Нидерланды |
| 51 | Seminar2018 22 марта 2018 — 16:00 Пунктуальное склеивание т-структур и комплекс мотивных пересечений разновидностей шимура. Доктор Вайбхав Вайш (ISI Bangalore). |
| 50 | Seminar2018 22 марта 2018 г. — 11:00 О гипотезах Тесье для множественности идеалов Проф. Дж. К. Верма (ИИТ Бомбей) |
| 49 | Лекция 2018 20 марта 2018 г. — 18:00 Ускоряющаяся расширяющаяся Вселенная: темная материя, темная энергия и космологическая постоянная Эйнштейна Профессор Бхарат Ратра (Университет штата Канзас) |
| 48 | Seminar2018 20 марта 2018 — 15:00 Темная энергия: постоянная или временная переменная? (и другие открытые вопросы) Профессор Бхарат Ратра (Университет штата Канзас) |
| 47 | Lecture2018 17 марта 2018 г. — 11:00 Специальная лекция о том, как машины обучаются? Доктор.Сарабджот Сингх Ананд |
| 46 | Seminar2018 16 марта 2018 — 17:00 Как неправильно складывается прионный белок Д-р Ишита Сенгупта, Национальный центр биологических наук |
| 45 | Seminar2018 16 марта 2018 — 17:00 Явная abc-гипотеза Бейкера Т. Н. Шори (ИИТ Бомбей) |
| 44 | Seminar2018 16 марта 2018 — 16:00 Гидрогеохимические процессы, их эволюция и перспективы на будущее Dr.Венкатраманан Сенапати, Школа системы опасностей для окружающей среды Земли, Национальный университет Пукён, Южная Корея, |
| 43 | Seminar2018 15 марта 2018 — 17:00 Функциональные наноматериалы: основы и приложения Д-р Субхра Яна, факультет DST INSPIRE, отделение химических, биологических и макромолекулярных наук, Национальный центр фундаментальных наук им. С. Н. Боса, Калькутта |
| 42 | Seminar2018 15 марта 2018 — 17:00 Прогнозирование закономерностей пространственно-временного распространения сигнала в сложных сетях Chittaranjan Hens, Dept.* -алгебры Д-р Рохит Д. Холкар (IISER Pune) |
| 40 | Seminar2018 12 марта 2018 — 17:00 Рассказ о двух городах: гравитация и термодинамика Доктор Суманта Чакраборти (МАКО, Калькутта) |
| 39 | Семинар 2018 12 марта 2018 — 16:00 Геохимические и изотопные подходы к изучению происхождения отложений на Индийском субконтиненте Д-р Нирадж Авашти, Отдел наук о Земле, Индийский технологический институт Канпур |
| 38 | Seminar2018 09 марта 2018 — 16:00 Помимо уравнения мелкой воды — конвективная параметризация и не только Dr.Бидют Бикаш Госвами, Центр прототипа климатического моделирования (CPCM), Нью-Йоркский университет, Абу-Даби, |
| 37 | Seminar2018 08 марта 2018 — 17:15 Комплексы бора переходных металлов: от основ к применению Проф. Сундаргопал Гош, ИИТ Мадрас |
| 36 | Seminar2018 7 марта 2018 — 17:00 Катализируемая миоглобином реакция внедрения карбена Д-р Викас Тьяги, ассистент. Профессор школы химии и биохимии, Технический и технологический институт Тапара, Патиала. |
| 35 | Seminar2018 05 марта 2018 — 17:30 Отражения искусства в скульптурах храмов Хойсала с особым упором на Натьяшастру и Нада-йогу Д-р Рохита Эсвер, кафедра древней истории, археологии, археологии и музеологии, Колледж Махараджаса, Университет Майсур |
| 34 | Seminar2018 05 марта 2018 — 17:30 Отражения искусства в скульптурах храмов Хойсала с особым акцентом на Натьяшастру и Нада-йогу Dr.Рохита Эсвер, факультет древней истории, археологии, археологии и музеологии, Колледж Махарараджа, Майсурский университет |
| 33 | Seminar2018 05 марта 2018 — 17:15 Разработка новых синтетических стратегий и тактик: их влияние, последствия и приложения Проф. Самбасиварао Котха, ИИТ Бомбей |
| 32 | Seminar2018 05 марта 2018 — 17:00 Квантово-оптический анализ в поисках квантовой теории, совместимой с некоммутативным подходом квантовой гравитации Dr.Санджиб Дей (IISER, Мохали) |
| 31 | Seminar2018 3 марта 2018 г. — 16:00 Развитие храмовой архитектуры в Южной Индии Д-р Читра Мадхаван; Фонд Калакшетры |
| 30 | Seminar2018 01 марта 2018 — 17:00 Самосборка анизотропных коллоидов Доктор Антара Пал, Отдел физической химии, Лундский университет, Лунд, Швеция |
| 29 | Seminar2018 28 февраля 2018 г. — 17:00 Взаимодействия световой материи и их ответы Dr.Пралок Кумар Саманта, Центр перспективных научных исследований Джавахарлала Неру, Бангалор |
| 28 | Seminar2018 27 фев 2018 — 16:00 Рассказы Джатаки: Путешествие в Древнюю Индию Латашри К.С.: общественный археолог; Основатель Chai With History; Инструктор курса, Университет Азима Премжи |
| 27 | Семинар 2018 26 фев 2018 — 17:00 Возвращение к функции массы HI Д-р Нишиканта Кхандаи (НИСЭР, Бхубанешвар) |
| 26 | Seminar2018 23 февраля 2018 г. — 17:05 Водородная связь и радиусы Ван-дер-Ваальса Проф.Эланганнан Арунан, IISC Bangalore |
| 25 | Семинар 2018 23 февраля 2018 г. — 15:00 Молекулярный механизм устойчивости к артемизинину у Plasmodium falciparum malaria Д-р Сувик Бхаттачарджи, Школа молекулярной медицины, JNU, Нью-Дели |
| 24 | Семинар 2018 22 февраля 2018 — 17:00 Фундаментальное понимание и применение химии иминоборонатов Д-р Анупам Бандиопадхьяй, Департамент химии, MIT-Кембридж |
| 23 | Seminar2018 22 фев 2018 — 17:00 Группы с инволюциями — их место в геометрии и теории представлений Проф.Дипендра Прасад (TIFR Mumbai) |
| 22 | Семинар 2018 19 фев 2018 — 16:00 Органический синтез, ориентированный на приложения: усилия нашей группы в области полного синтеза и медицинской химии Д-р Шриниваса Редди, CSIR-NCL Пуна |
| 21 | Семинар 2018 16 фев 2018 — 17:00 Механохимическое упрочнение полимерных материалов с помощью пьезоэлектрических наночастиц Д-р Хемакеш Мохапатра, Институт молекулярной инженерии, Чикагский университет |
| 20 | Seminar2018 16 февраля 2018 — 16:00 Оптимизация связывания углерода за счет увеличения нефтеотдачи CO2 в зрелом индийском пласте: комплексный подход Dr.Шиб Санкар Гангули, факультет DST Inspire, IIT Kanpur |
| 19 | Seminar2018 16 фев 2018 — 15:00 Здоровое старение: от гомеодинамики к гормезису Проф. Суреш Раттан, доктор философии, доктор наук, Лаборатория клеточного старения, Департамент молекулярной биологии и генетики; Орхусский университет, Дания |
| 18 | Конференция / семинар 2018 16 февраля 2018 г. — 08:00 24-я конференция Национального общества магнитного резонанса Индии |
| 17 | Seminar2018 15 февраля 2018 г. — 17:00 Настройка поверхности наночастиц: от сборки, индуцированной кластером, к динамике на уровне отдельных частиц Dr.Индранат Чакраборти, научный сотрудник Александра фон Гумбольдта и младший руководитель группы, Центр гибридной наноструктуры (CHyN), Гамбургский университет |
| 16 | Seminar2018 13 февраля 2018 — 16:00 Трехмерная магнитотеллурическая визуализация: истории развития тектонической эволюции систем «Металлогенические провинции» и «Нефтяной бассейн» с двух разных континентов Д-р Вед Пракаш Маурья, Координатор геофизики (COGE) ) в Observatorio Nacional (ON), Рио-де-Жанейро, Бразилия |
| 15 | Colloquia2018 13 февраля 2018 — 16:00 Квантовая теория точна или приблизительна? Проф.Теджиндер П. Сингх, Институт фундаментальных исследований Таты, Мумбаи |
| 14 | Конференция / семинар 2018 10 февраля 2018 г. — 09:00 Семинар по мягким навыкам Д-р В. В. С. Шарма и г-н Муралидхар Дасьяпу (Центр управления отношением посредством образования и обучения) |
| 13 | Seminar2018 07 февраля 2018 — 10:00 Открытие, структура и спектроскопия литических полисахаридных монооксигеназ: ферменты для изменения мира биотоплива Проф. Пол Уолтон, Йоркский университет, Великобритания |
| 12 | Seminar2018 02 фев 2018 — 16:00 Технологические и научные достижения во время Хараппанской цивилизации Проф.Васант Шинде (Колледж Декан, Пуна) |
| 11 | Seminar2018 01 февраля 2018 — 05:15 Устойчивые подходы в медицинской химии: новые концепции и приложения Проф. Асит К. Чакраборти, NIPER Mohali |
| 10 | Colloquia2018 31 января 2018 — 16:00 Редактирование генома in vivo для наследственных заболеваний печени Проф. Джаянта Рой-Чоудхури, Медицинский колледж Альберта Эйнштейна, Бронкс, Нью-Йорк, США |
| 9 | Seminar2018 23 января 2018 — 15:00 Клеточная геометрия регулирует ответ на передачу сигналов TNFα Dr.Анинда Митра, Институт механобиологии, Национальный университет Сингапура, Сингапур, и Институт молекулярной онкологии FIRC (IFOM), Милан, Италия |
| 8 | Seminar2018 22 января 2018 — 17:00 Применение кремниевых сенсоров в физике высоких энергий и астрономии Проф. Шашикант Р. Дугад (TIFR, Мумбаи) |
| 7 | Семинар 2018 15 января 2018 — 17:00 Граничные условия в гравитации Д-р Кришна Мохан Паратту (ИИТ Мадрас) |
| 6 | Seminar2018 15 января 2018 — 15:00 Генетические основы естественной изменчивости гомеостаза железа и роста корней Dr.Сантош Б. Сатбхай, Институт биологических исследований Солка, Сан-Диего, США |
| 5 | Seminar2018 12 января 2018 г. — 17:15 Хиральные N-сульфинилимины: новые открытия Проф. Мигель Юс, кафедра органической химии, Университет Аликанте, Испания. |
| 4 | Seminar2018 12 января 2018 — 16:00 Металлические комплексы чеканки как хиральные катализаторы для асимметричного 1,3-диполярного циклоприсоединения азометина илида Профессор Кармен Нагера, факультет органической химии, Университет Аликанте, Испания. |
| 3 | Seminar2018 09 января 2018 — 16:00 Земля и труд на «границах» Калькутты: беженец живет между городом и страной Доктор Химадри Чаттерджи, научный сотрудник, Департамент гуманитарных и социальных наук, IISER Mohali |
| 2 | Seminar2018 05 января 2018 — 17:00 Группы с двумя размерами классов сопряженности и классом нильпотентности 3 Доктор Рахул Киттюр |
| 1 | Семинар 2018 02 января 2018 — 15:00 Исследование клеточного цикла отдельных клеток Dr.Кедар Натараджан, Европейский институт биоинформатики, Кембридж, Великобритания |
Синергетическое смешение ценных гетероциклов подавляет рост Plasmodium falciparum в культуре и инфекцию P. berghei в модели мыши.
Дизайн и синтез соединения
Мы разработали химическую стратегию, которая способствовала ценному слиянию Pht, бензимидазола и триазола. Были предприняты многочисленные попытки изменения триазольного каркаса, включая замещенные ароматические кольца, алкильные цепи и гидрофильные заместители для улучшения профиля активности.В качестве линкеров использовали аминокислоты с алифатическими цепями, то есть валин, лейцин и изолейцин. Был разработан простой и рационально совместимый синтетический маршрут (рис. 1) для создания библиотеки новых аналогов Pht. Синтез связанного бензимдазолом Pht, ключевого синтона, начался с сочетания N -замещенного изоиндолин-1,3-диона и o -фенилендиамина, 2 с последующей катализируемой уксусной кислотой циклизацией при температуре кипения с образованием промежуточных продуктов 3a – c .Промежуточное соединение алкина 5a получали алкилированием 3a пропаргилбромидом ( 4 ). Первоначально соединение 5a показало выход только 20%, однако попытки стандартизировать условия реакции привели к выходу 62%. Аналогичным образом соединения 5b и 5c были выделены с выходами 47% и 72% соответственно.
Рисунок 1Синтез аналогов Pht 6a — e ‘.
Впоследствии алкилированные продукты 5a – c были обработаны различными азидами (реакции щелчка) под воздействием микроволнового излучения для получения конечных продуктов 6a — 6e ′.Объем различных азидов для синтеза новых аналогов Pht показан в таблице S1. Все синтезированные аналоги Pht были охарактеризованы различными спектроскопическими методами ( 1 H ЯМР, 13 C ЯМР, HRMS и ИК-спектроскопия и т. Д.). Кроме того, монокристаллы были выращены для одного репрезентативного аналога 6a и подвергнуты дифракции рентгеновских лучей на монокристаллах. Молекулярная диаграмма 6a изображена на рис. 2.
Рис. 2Молекулярная структура 6a с уровнем вероятности 40%.
Соединение 6a кристаллизовалось в моноклинной системе с пространственной группой C2 / c . Подробности сбора данных, решения структуры и уточнения перечислены в таблице S2.
Биологические исследования и анализ взаимосвязи структура-активность (SARs)
Антималярийная активность всех перечисленных аналогов Pht была первоначально оценена на асинхронных культурах клона P. falciparum 3D7 ( Pf 3D7) с использованием теста SYBR Green.Наиболее активными были соединения со средней концентрацией 50% ингибирования роста (IC 50 ) <5,14 мкМ, установленной в качестве контрольного порогового значения IC 50 на основе среднего значения эталонной молекулы Pht ( I ) IC 50 Средние значения IC 50 основаны на трех отдельных экспериментах (таблица 1). Четыре новых аналога 6a , 6 h , 6 m и 6 u проявили антиплазмодиальную активность против штамма Pf 3D7 с IC 50 <5.14 мкМ. Кроме того, все четыре соединения ингибировали рост устойчивого к CQ штамма Pf W2 с концентрациями IC 50 ниже контрольного порогового значения IC 50 ; 6a IC 50 = 0,7 (± 0,01) мкМ, 6 ч IC 50 = 1,3 (± 0,11) мкМ, 6 м IC 50 = 3,8 (± 0,34) мкМ и 6 мкМ IC 50 = 0,9 (± 0,6) мкМ. Двенадцать из 31 аналога Pht не обладали дозозависимым действием на рост паразитов, следовательно, IC 50 был недостижим (без ответа на дозу, NDR).CQ и DHA были протестированы вместе с Phts для обеспечения качества и контроля анализа, и значения IC 50 , полученные SYBR Green, для обоих стандартных противомалярийных средств были в соответствии с рекомендациями литературы 30 .
Таблица 1 SAR-исследование заместителя R ‘: ароматические кольца. Примечание: Иллюстрация аналогов Pht, Pht (I), а также значений IC 50 хлорохина и дигидроартемизинина.Взаимосвязь структура-активность сосредоточена на различных заменах триазольного кольца и аминокислот (таблицы 1 и 2).Функционализированные Phts были синтезированы и подвергнуты скринингу на противомалярийную активность в отношении культивированного Pf 3D7 и сравнены с незамещенным Pht I (IC 50 = 5,14 ± 1,67 мкМ), CQ (IC 50 = 0,03 ± 0,67 мкМ) и DHA ( IC 50 = 0,003 ± 0,3 мкМ). При конструировании новых молекул Pht, бензимидазольные каркасы оставались неизменными, а его фланкирующие стороны, триазолы, были диверсифицированы. Из результатов скрининга (таблицы 1 и 2) очевидно, что вариации заместителей R или R ‘играют важную роль в эффективности соединений против роста паразита.
Таблица 2 SAR противомалярийной активности заместителя R ‘: алифатические и гликозидные группы.Противомалярийные данные показывают, что введение более крупных групп R (изобутила или сек, -бутил) увеличивает эффективность, как это было замечено в случае Phts 6a , 6h и 6u , тогда как относительно меньшие группы снижают активность (т.е. 6м ). Как показано в таблице 1, введение фенильного заместителя также влияет на эффективность молекул.Аналоги Pht, содержащие 4-фторфенильное кольцо в триазольном фрагменте с R, представленным как бутильная группа ( 6h ), были замечены как более активные по сравнению с ананлогами, содержащими замещенную фенильную группу в триазольном фрагменте. Этот результат, по крайней мере частично, по-видимому, связан с высокой электроотрицательностью фторгруппы. Аналоги с незамещенным ароматическим кольцом также проявляли значительное ингибирование роста паразита, но только тогда, когда R был заменен изобутильной группой (например, 6a ).В отсутствие функциональной группы в триазольном кольце с R, представленным как изобутильная группа, мы наблюдали самую высокую эффективность против штамма P. falciparum 3D7 (т.е. 6u ).
Стадия и влияние на титры паразитемииЗатем мы попытались определить стадийную специфичность антиплазмодиальной активности четырех активных аналогов ( 6a , 6h , 6m и 6u ) на синхронизированных культурах штамма Pf 3D7 при гематокрите 2% и 1% паразитемия с концентрациями, соответствующими индивидуальному препарату IC 50 .Чтобы определить влияние соединений на паразитов как на ранней, так и на поздней стадии цикла, обработку проводили на недавно синхронизированных кольцах в течение 12 часов, а влияние лекарственного средства на рост и морфологию паразитов контролировали через 6 и 12 часов после воздействия. Точно так же первые трофозоиты подвергались воздействию каждого соединения и инкубировались в течение 16 часов, а затем контролировались через 6 и 16 часов после воздействия. Влияние соединений на морфологию и развитие паразитов сравнивали в лунках с лекарственными средствами, подвергнутыми воздействию, и без них (рис. 3 и 4).Соединения 6a и 6u были активны против форм на стадии кольца, на что указывает заметное снижение плотности паразитов и аномальная морфология стадии кольца через 6 часов и 12 часов после воздействия. Однако соединения 6a и 6u не препятствовали развитию паразитарной прогрессии от трофозоитов к шизонтам при воздействии на них отдельной культуры трофозоитов. Обработка 6h и 6m не останавливала созревание на стадии кольца, на что указывает заметное увеличение роста на стадии кольца через 12 часов, как в группе без обработки.Однако при исследовании как 6 часов , так и 6 часов на зрелых стадиях крови, 6 часов приводили к полному разрушению трофозоитов через 6 часов после воздействия лекарственного средства на трофозоиты на ранних стадиях. Кроме того, наблюдение за эффектом 6ч через 16 часов показало присутствие шизонтов с аномальной морфологией.
Рисунок 3Микрофотографии P. falciparum кольцевых форм, обработанных аналогами Pht. Иллюстрация влияния лечения аналогами Pht на раннюю стадию эритроцитарного паразита (кольца).(Все процедуры выполнялись параллельно с группой без лечения).
Рисунок 4Микрофотографии стадий трофозоита P. falciparum , обработанных аналогами Pht. Примечание: Иллюстрация влияния лечения аналогами Pht на раннюю (кольца) и зрелую (трофозоиты и шизонты) стадии крови паразитов. (Все процедуры выполнялись параллельно с группой без лечения).
Хотя на рост трофозоитов, по-видимому, не влияла обработка 6m через 6 часов, полученные шизонты выглядели менее зернистыми и не имели различимых мерозоитов после 16 часов воздействия.Влияние аналогов на количество паразитозов коррелировало с их стадийной специфичностью. Аналоги 6a и 6u вызывали снижение паразитемии на стадии кольца через 6 часов после воздействия, в то время как их влияние на титры паразитов стадии зрелой крови через 16 часов было незначительным (рис. 5). Хотя 6h и 6m не влияли на рост паразитов через 6 часов после воздействия (т. Е. На стадии кольца), оба аналога вызывали снижение паразитемии через 16 часов (т.5).
Рисунок 5Влияние аналогов Pht на титры роста паразитов (Примечание: графическое описание ингибирующего действия выбранных аналогов на титры роста паразитов через 6 часов после инкубации с кольцевыми стадиями и через 16 часов после инкубации лекарств с трофозоитной стадией. Процент паразитемии был получен путем подсчета числа инфицированных эритроцитов из 2000 эритроцитов на тонких мазках, окрашенных по Гимзе, из каждого эксперимента. Столбчатые диаграммы представляют собой среднее значение трех различных экспериментов).
Анализы лекарственного взаимодействия
Затем мы исследовали синергетическую ингибирующую активность лекарств между аналогами Pht и CQ или DHA. Синергетическая ингибирующая активность наблюдалась между тремя аналогами ( 6a , 6h и 6u ) в сочетании как с CQ, так и с DHA против штаммов 3D7 и W2 (Таблица S3). Определение лекарственных взаимодействий между аналогами с CQ и DHA необходимо для выявления возможных лекарственных препаратов-партнеров для борьбы с устойчивостью к современным противомалярийным препаратам.Недавние сообщения о появлении чувствительных к CQ штаммов P. falciparum , достигнутые в некоторых эндемичных по малярии регионах, приписывают снятию давления CQ на популяции паразитов из-за замены сульфадоксин-пириметамином (SP) и ACT 31, 32 . CQ был идеальным противомалярийным средством из-за его фармакокинетики, профиля безопасности и низкой стоимости. Поскольку устойчивость к P. falciparum развивалась в первую очередь из-за введения в качестве монотерапии, определение потенциальных лекарственных препаратов-партнеров CQ может способствовать снижению развития исторической устойчивости паразитов к монотерапии CQ.Артемизинин (АРТ) и его производные в настоящее время являются передовой линией защиты от неосложненной малярии P. falciparum и применяются как АКТ. Появление устойчивости к обоим компонентам препарата АКТ требует идентификации заместительной АРТ и возможных комбинационных хемотипов 33,34,35 . Примечательно, что три аналога Pht ( 6a , 6h и 6u ) показали синергетическую активность в сочетании с CQ и DHA против Pf 3D7 и Pf W2, что указывает на их потенциал для использования в комбинированной терапии.
Противомалярийный эффект только аналогов Pht и в комбинации с артемизинином у мышей, инфицированных Plasmodium berghei
Антималярийный эффект двух активных аналогов, 6h и 6u (вводимых в дозе 50 мг / кг массы тела), был определен в мыши, инфицированные штаммом P. berghei NK65, который приводит к высокому уровню паразитемии на стадии крови. Введение либо 6h , либо 6u отдельно в течение четырех дней подряд вызывало подавление паразитарной нагрузки на 5 и 8 дни инфекции и улучшало выживаемость по сравнению с нелеченными (рис.6А и В). Однако аналог 6u имел лучшую противомалярийную эффективность, чем аналог 6h (рис. 6А). Таким образом, затем мы оценили эффективность 6u (50 мг / кг) в комбинации с артемизинином (5 мг / кг веса тела) в уменьшенной дозировке на модели малярии на мышах. Как показано на фиг.6, ни одно из соединений, доставляемых в виде монотерапии, не приводило к избавлению от паразитемии, однако совместное введение аналога 6u с АРТ значительно повышало противомалярийную эффективность за счет снижения паразитарной нагрузки и увеличения выживаемости ( P < 0.05, рис. 6А и В). Среднее время выживания животных, получавших только Pht 6u , а также в комбинации с АРТ, составляло 23 и 27,5 дня соответственно ( P <0,001). Эти результаты демонстрируют, что соединение 6u в сочетании с АРТ имеет наибольшую терапевтическую эффективность на мышиной модели малярии.
Рисунок 6Противомалярийный эффект аналогов Pht ( 6h и 6u ) отдельно и в комбинации с артемизинином ( 6u и ART) на паразитемию и выживаемость мышей, инфицированных P.berghei NK65. Мышам вводили 1 × 10 7 инфицированных P. berghei эритроцитов. Через 48 часов мышам вводили контроль (PBS : , необработанный ), 6 часов и 6u (отдельно) и 6u в сочетании с артемизинином (все вводили подкожно) в течение четырех дней подряд. % Паразитемии определяли для групп на 5, 8 и 15 дни путем случайного выбора 10 различных оптических полей на мазках крови. ( A ) Администрирование 6h и 6u (отдельно) и 6u в сочетании с ART.Данные представляют собой среднее значение ± стандартная ошибка среднего для шести животных в каждой экспериментальной группе. ( B ) Выживаемость в группах лечения (n = 6 / группа). ( C ) Микрофотография мазков крови необработанных групп по сравнению с группами лечения на 15-й день после инфицирования (100-кратное увеличение). АРТ, Артемизинин; * P <0,05.
Оценка цитотоксичности
На последнем этапе исследований мы определили цитотоксический эффект четырех активных аналогов ( 6a , 6h , 6m и 6u ) в клеточных линиях U937 путем измерения 50% цитотоксичности. концентрации (CC 50 ) и расчет индексов селективности (SI) в качестве меры токсичности для клеток человека.
Соединения 6a и 6m показали сильные значения CC 50 <1 мкМ, 0,91 ± 0,32 мкМ и 0,78 ± 0,32 мкМ, соответственно, и считались обладающими более высокой селективностью для клеточных линий U937 по сравнению с P. falciparum (значения SI 1,01 ± 1,5 мкМ и 1,11 ± 1,5 мкМ соответственно). Аналоги 6h и 6u обладали меньшей селективностью в отношении линий клеток U937 со значениями CC 50 28,82 ± 0,67 мкМ и 2,08 ± 1,6 мкМ соответственно и значениями SI 41.2 ± 1,7 мкМ и 2,31 ± 0,76 мкМ, соответственно, и поэтому считались менее токсичными для клеток человека. Таким образом, дополнительные химические модификации 6a и 6m могут потребоваться для получения аналогов с меньшей селективностью и токсичностью для линий клеток человека в контексте сохранения их антиплазмодной активности.
Аутоиммунитет и оптимальный витамин D — стохастическая химическая динамическая корреляция — июнь 2014 г.
Стохастический химический динамический подход для корреляции аутоиммунитета и оптимального диапазона витамина D
Сусмита Рой, Кришна Шринивас, Биман Багчи profbiman в Gmail.com
SSCU, Индийский институт науки, Бангалор, Карнатака, Индия
Фигурка F4 (снизу)
На основании нескольких недавних экспериментальных наблюдений, что витамин D может взаимодействовать с антигенпрезентирующими клетками (APC) и Т-лимфоцитарными клетками (Т-клетками), чтобы стимулировать и регулировать различные стадии иммунного ответа, мы разработали грубозернистую, но общую кинетическую модель. в попытке уловить роль витамина D в иммуномодулирующих реакциях.Наша кинетическая модель, разработанная с использованием идей теории химических сетей, приводит к системе из девяти связанных уравнений, которые мы решаем как прямым, так и стохастическим (Гиллеспи) методами. Оба анализа последовательно предоставляют подробную информацию о зависимости иммунного ответа от изменения параметров критической скорости. Мы обнаружили, что, хотя витамин D играет незначительную роль в начальном иммунном ответе, он оказывает сильное влияние в долгосрочной перспективе, особенно помогая системе достичь нового, стабильного устойчивого состояния.В исследовании исследуется роль витамина D в сохранении наблюдаемой бистабильности на фазовой диаграмме (охватываемой системными параметрами) иммунной регуляции, что позволяет ответу выдерживать широкий спектр патогенных стимуляций, которые могут помочь в сопротивлении аутоиммунным заболеваниям. Мы также изучаем, как витамин D влияет на зависящую от времени популяцию дендритных клеток, которые связаны между врожденными и адаптивными иммунными ответами. Вариации дозозависимой реакции противовоспалительных и провоспалительных популяций Т-клеток на витамин D хорошо коррелируют с недавними экспериментальными результатами.Наша кинетическая модель позволяет оценить диапазон оптимального уровня витамина D, необходимого для бесперебойного функционирования иммунной системы и для контроля как гиперрегуляции, так и воспаления. Что наиболее важно, настоящее исследование показывает, что передозировка или токсический уровень витамина D или любого стероидного аналога может вызвать слишком сильный толерантный ответ, что приведет к неэффективности адаптивной иммунной функции.
Образец цитирования: Рой С., Шринивас К., Багчи Б. (2014) Стохастический химический динамический подход для корреляции аутоиммунитета и оптимального диапазона витамина D.PLoS ONE 9 (6): e100635. DOI: 10.1371 / journal.pone.0100635
Редактор: Энрике Эрнандес-Лемус, Национальный институт геномной медицины, Мексика Получено 16 декабря 2013 г .; Принята в печать 29 мая 2014 г .; Опубликовано 27 июня 2014 г.
Авторские права: © 2014 Roy et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа была частично поддержана грантами Министерства науки и технологий (DST) Индии и сэра Дж.C. Bose Fellowship (DST). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Ниже приведены лишь некоторые рисунки и уравнения в PDF-документе .
Введение
Сообщается, что витамин D участвует в большом количестве различных иммунных ответов [1-6], хотя наше количественное понимание этих процессов на клеточном уровне все еще остается в значительной степени неполным.Это связано с огромной сложностью иммунной системы человека, которая зависит от большого количества взаимодействующих (некоторые из них могут быть еще неизвестны) компонентов. Кроме того, иммунная система в целом делится на две ветви: врожденный иммунитет и адаптивный иммунитет. В то время как первая ветвь является универсальной по действию, последняя весьма специфична. Вдохновленные современными эпидемиологическими исследованиями, в последние два десятилетия усилия были направлены на понимание происхождения неклассических иммуномодулирующих реакций, которые, как полагают, запускаются активным 1,25-дигидроксивитамином-D [1-6].Помимо установленной классической функции в метаболизме кальция, исследования витамина D в настоящее время все больше фокусируются на его плейотропном действии [1-6].
Иммунотерапия, опосредованная витамином D, наблюдалась в течение последних 150 лет. С начала 1900-х годов жир печени трески и УФ-свет стали широко признаны в качестве основных источников витамина D. Терапевтическое использование витамина D впервые привлекло внимание в 1849 году, когда доктор Чарльз Джеймс Блазиус Уильям использовал рыбий жир для лечения более 400 больных туберкулезом [7].После долгого 50-летнего перерыва Нильс Финсен получил Нобелевскую премию, подчеркнув лечебную ценность ультрафиолетового излучения, с помощью которого он лечил более 800 пациентов, страдающих волчанкой (кожная форма ТБ) [8,9]. В индийских традиционных аюрведических методах лечения использование солнечного света для лечения и уменьшения болезней восходит к нескольким тысячам лет назад (), где его называют «Сурьявиган» (Значение: наука о солнечном свете).
Витамин D играет разные роли как во врожденном, так и во адаптивном иммунитете. Несколько экспериментальных и клинических исследований показали, что эндогенно продуцируемый активный витамин D (1,25 (OH) 2D3) в макрофагах увеличивает скорость производства антимикробных пептидов (кателицидина, b-дефенсинов и т. Д.), Способствуя развитию врожденного иммунитета [10 , 11].Впоследствии превращение 25-D3 в функциональный 1,25-D3 (известный как активный витамин-D) в антигенпрезентирующих клетках (APC, таких как дендритные клетки, макрофаги) оказывает сильное влияние на адаптивную иммунную систему [12]. Предыдущие эпидемиологические данные подчеркивают связь между недостаточностью витамина D и рядом иммуно-опосредованных заболеваний, а именно различных типов аутоиммунных заболеваний. Экспериментальные исследования иммуномодулирующих свойств витамина D показывают, что аутоиммунитет в первую очередь обусловлен увеличением количества Т-хелперных клеток (например,грамм. Th2), которые атакуют различные ткани тела. В частности, ингибирующее действие витамина D на такие провоспалительные Т-клеточные ответы и стимулирование регуляторных Т-клеток (TReg) может, по крайней мере частично, объяснить некоторые из этих ассоциаций [11-15].
Некоторые недавние экспериментальные исследования пролили свет на такие регуляторные действия, проявляемые как витамином D, так и регуляторными Т-клетками, и их взаимодействие в сопротивлении аутоиммунитету. Различные функции эффекторных Т-клеток (кратко описанные в тексте S1 в файле S1) [16], как часто обнаруживают, развиваются в присутствии антигена, процессируемого и представляемого антигенпрезентирующими клетками (APC), которые побуждают соответствующие костимуляторные сигналы к индуцируют созревание наивных Т-клеток [17,18].В 2000 году Jonuleit с соавторами охарактеризовали различные типы Т-клеточных ответов, которые критически зависят от фазы созревания дендритных клеток (ДК) [19]. Они сообщили, что в то время как стимуляция зрелыми DCs способствует пролиферации воспалительных Th2-клеток, контакты наивных Т-клеток с незрелыми DC индуцируют IL-10, продуцирующий регуляторные 1-подобные ответы Т-клеток [19]. В 2003 г. Powrie и Maloy предложили схему взаимодействия, объясняющую такую взаимосвязь между APC и Т-клеточными ответами [20].В тот же период времени Piemonti и соавторы упомянули об особой роли 1,25 (OH) 2D3 в модуляции иммунных ответов посредством ингибирования дифференцировки DC и созревания в мощные APC [21]. Активная форма витамина D отрицательно влияет на активацию, пролиферацию и дифференцировку Т-клеток, одновременно способствуя выработке регуляторных Т-клеток (TReg), которые функционируют как эффективный иммунный регулятор [22-24]. Недавно Correale et al. показали, что эффекторные Т-клетки способны метаболизировать неактивный 25 (ОН) D3 в биологически активный 1,25 (ОН) 2D3, поскольку эти Т-клетки экспрессируют фермент 1 -гидроксилазу, который конститутивно способствует этому превращению [25].
В настоящем исследовании мы разрабатываем теоретическую модель крупнозернистой кинетической сети на основе вышеупомянутых экспериментальных наблюдений. Наша основная цель — количественно изучить зависимость иммунитета от витамина D и изучить его возможную роль в снижении риска аутоиммунных заболеваний и смертельных инфекций. Мы анализируем несколько иммуномодулирующих реакций, которые контролируются витамином D, включая как врожденные, так и адаптивные ответы, как указано в нескольких экспериментальных отчетах и обзорах [11–12, 26].Хотя существует множество сложных биохимических реакций и задействованных реагентов, мы рассмотрели только определенное их количество, которые являются важными компонентами иммунной системы и имеют прямое взаимодействие с витамином D.
Мы решаем проблему оптимального диапазона потребления витамина D, которая была поднята международным агентством Всемирной организации здравоохранения по исследованиям рака. Настоящее исследование предполагает, что ингибирующее действие, оказываемое регуляторными Т-клетками, индуцированными витамином-D и самим витамином-D, на ответ эффекторных Т-клеток, может играть важную роль в предотвращении аутоиммунных заболеваний.
Несколько ранних математических моделей также изучали воспалительную роль эффекторных Т-клеток и их регуляцию регуляторными Т-клетками. В последние годы Friedman et al. изучили влияние Т-клеток на воспалительное заболевание кишечника [27]. Pillis с соавторами исследовали влияние регуляторных Т-клеток на лечение почечно-клеточного рака [28]. В другой модели Villoslada et al. наблюдали динамическую перекрестную регуляцию антигенспецифических эффекторных и регуляторных субпопуляций Т-клеток в связи с микроглией при аутоиммунной реакции мозга [29].Пожалуй, наиболее актуальная модель регуляции Т-клеток в иммунной системе была представлена в недавней статье Fouchet et al. [30]. Они определили важные ингредиенты иммунной системы и сформулировали связанные уравнения скорости для всего процесса, чтобы показать регуляцию эффекторных и регуляторных Т-клеток путем изменения различных констант скорости.
Хотя все эти модели довольно аккуратны, они не учитывают основные эффекты витамина D [27-30], тогда как несколько экспериментов уже показали важность витамина D как для врожденной, так и для адаптивной иммунной системы.Здесь мы имплантировали нелинейный эффект витамина D в базовую модель регуляции Т-клеток. Нелинейный эффект витамина D является косвенным результатом презентации антигена и последующего образования эффекторных Т-клеток. Производство эффекторных Т-клеток сигнализирует об активации витамина D, который, в свою очередь, подавляет продукцию эффекторных Т-клеток. Эта модель в первую очередь стремится понять активацию ответов Т-клеток и влияние витамина D на толерантность / регуляторную природу этих ответов.Поэтому мы подчеркнули регулирующую функцию витамина D в адаптивной иммунной системе. Мы предположили, что врожденный механизм уничтожает патогены с постоянной скоростью, продуцируя некоторые антимикробные пептиды, и это приводит к управлению первичной защитой от инфекционных заболеваний.
Важными составляющими модели, рассматриваемой здесь, являются следующие: (i) патоген (важно отметить, что в нашем анализе мы рассматривали патоген как числовую величину P , которая способна вызывать опосредованную Т-клетками иммунную ответ), (ii) наивные Т-клетки, (iii) миелоидные дендритные клетки в форме профессиональных антигенпрезентирующих клеток (APC), как в их покоящихся (незрелых), так и в активированных (зрелых) формах (iv) эффекторных и регуляторных Т-клетках. клетки, (v) неактивный витамин D (25 (OH) D3) и активный витамин D (1,25 (OH) 2D3).Однако участники, такие как рецептор витамина D (VDR) и фермент 25 (OH) D3-1a-гидроксилаза (CYP27B1), которые одновременно превращают неактивный витамин D в активный витамин D (1,25 (OH) 2 D) -VDR [D * -VDR] белковый комплекс, рассматриваются как неявные факторы для активации необходимого транскрипционного мотива.
Здесь важно подчеркнуть, что мы по существу объединили три важных экспериментальных наблюдения, которые включают основные особенности адаптивных ответов, о которых сообщили (i) Powri и Maloy, [20] (ii) Jorge Correale et al.[25] и (iii) Лоренцо Пьемонти и др. [21]. На рисунке 1 мы представили модель сложной сети взаимодействия, которая включает в себя различные компоненты, их взаимосвязь и регуляцию, задействованные в иммунной системе.
Настоящий подход к построению сетевой кинетической модели очень похож на аналогичные методы, принятые ранее при изучении кинетического считывания доказательств [31,32], а также в кинетике ферментов [3335]. Во всех этих исследованиях точному количественному прогнозированию препятствует недостаточное знание параметров системы; тем более что значения констант скорости часто недоступны.Этот пробел действительно является источником серьезной проблемы не только при изучении кинетической корректуры и кинетики ферментов, но также, как мы находим здесь, в теоретических исследованиях иммунологии. Наконец, основные уравнения, используемые во всех таких задачах, решаются с использованием метода среднего времени первого прохождения [32,36,37], алгоритма Гиллеспи или прямого численного интегрирования. Мы принимаем как детерминированный подход, решая дифференциальные уравнения численно, так и стохастическое моделирование с использованием алгоритма Гиллеспи [38,39].В разделе, посвященном методам, обсуждается крупнозернистая схема взаимодействия, разработка схемы реакции и основные уравнения. Однако все представленные здесь результаты оцениваются с помощью метода стохастического моделирования.
Значения задействованных параметров (константы скорости и концентрации) могут охватывать широкий диапазон и могут варьироваться от случая к случаю. Таким образом, было проведено исследование реакции на изменение важных параметров. Такое исследование явно необходимо в данном контексте.
Результаты
При патогенной атаке здоровая иммунная система реагирует повышением скорости пролиферации и дифференцировки эффекторных Т-клеток [40]. Однако недостаточное подавление генерации эффекторных Т-клеток часто может приводить к инициации аутоиммунитета, когда нарушается толерантность к аутоантигенам [41]. Такие события являются результатом слабой регуляции нашей иммунной системы, в которой эффекторные Т-клетки многочисленны, а уровни регуляторных Т-клеток довольно низкие.
Рисунок 1. Схематическое изображение адаптивных иммунных ответов с точки зрения клеточных взаимодействий, включая витамин D, основанное на некоторых экспериментальных результатах и клинических наблюдениях.
На схеме основными событиями являются следующие: (1) Главный шаг — уничтожение патогена эффекторными Т-клетками. (2) В присутствии патогена неактивные APC стимулируются после распознавания патогена и образуют покоящиеся APC. (3) Покоящиеся APC активируются либо патогеном, либо присутствием какой-либо эффекторной Т-клетки [19,20].(4) Активация эффекторных Т-клеток инициируется этими активными APC. (5) Эффекторные Т-клетки инициируют образование активного витамина D из его неактивной формы [24]. (6) Отдыхающие APC и активный витамин D могут стимулировать производство регуляторных Т-клеток из их предшественника [25]. (7) Повышение скорости продукции эффекторных Т-клеток контролируется как регуляторными Т-клетками (TReg), так и активным витамином D [24]. (8) Кроме того, витамин D и регуляторные Т-клетки активируют образование АРС в состоянии покоя из активных АРС [21].
Но здоровая иммунная система обычно функционирует со сбалансированной регуляцией, которая контролирует популяцию эффекторных Т-клеток до соответствующего уровня, который является адекватным для очистки от патогенов. Производство эффекторных Т-клеток снова зависит от процесса активации APC, контролируемого двумя параметрами скорости: скоростью активации APC патогенной стимуляцией (kinp) и скоростью активации APC эффекторными T-клетками (krese). Здесь появляется роль витамина D, оптимальный уровень которого эффективно поддерживает этот баланс в иммунной регуляции.Витамин-D эффективно стимулирует активность регуляторных Т-клеток. Более того, сам витамин D снижает гиперактивность APC и эффекторных Т-клеток. Напротив, иммунная система может также достичь строго регулируемого состояния, в котором эффекторные клетки сильно подавлены регулирующими эффектами витамина D и / или регуляторных Т-клеток.
В других модельных исследованиях предполагается, что только регуляторные Т-клетки поддерживают сбалансированную регуляцию [28-30]. Существует несколько экспериментальных и клинических наблюдений, показывающих важную роль витамина D и его зависимые от концентрации эффекты в иммунной регуляции.Однако мы не знаем ни одного теоретического модельного исследования, которое использовалось бы для изучения такой интересной роли витамина D.
Влияние витамина D на популяцию Т-клеток: переход от слабой регуляции к сильной
Противоположная роль регуляторных и эффекторных Т-клеток в иммунологической активности и их соответствующая регуляция витамином D часто определяют силу иммунной регуляции и конечную судьбу заболевания. Регуляция в значительной степени определяется активацией APC с последующей продукцией эффекторных Т-клеток.В настоящем исследовании мы разделили регуляцию на три группы на основе APC и параметра взаимодействия эффекторных Т-клеток (krese): (i) сильная регуляция, (ii) умеренная регуляция и (iii) слабая регуляция. Чтобы исследовать несколько факторов, связанных с витамином D, мы провели анализ временной эволюции каждого участвующего элемента после патогенной атаки, чтобы изучить их долгосрочное поведение. Мы изучили все три ограничения регуляции, варьируя крезу как в отсутствие, так и в присутствии витамина D при различных патогенных стимуляциях (кинп).
Численные результаты решения нашей системы уравнений показаны на Рисунке 2 в виде серии кривых для всех трех пределов регулирования, как в присутствии, так и в отсутствие витамина D. Результаты весьма показательны, и мы обсудим их более подробно ниже.
Из рисунка 2 (а) мы видим, что в отсутствие витамина D система попадает под строгий предел регулирования, когда мы фиксируем krese = 10. Для сравнения, наличие стандартного уровня витамина D при таком же пределе креза эффективно сохраняет эту сильную регуляцию (рис. 2 (b)).При krese = 102 мы находим бистабильную область, где как сильные, так и слабые регуляции сосуществуют как в отсутствие, так и в присутствии витамина D (рисунки 2 (c) и 2 (d)). Такое бистабильное поведение можно охарактеризовать как умеренную регуляцию Т-клеточного ответа. В одном из ранних исследований Fouchet и соавторы [30] проанализировали устойчивые значения Т-клеток в этих трех интервалах регуляции и показали похожие интересные явления, но в отсутствие витамина D. Когда мы сдвигаем интервал умеренной регуляции в сторону слабо регулируемого состояния (при krese = 103), присутствие стандартного уровня витамина D по-прежнему вызывает умеренную регуляцию стабильной популяции эффекторных Т-клеток по сравнению с слабая регуляция в отсутствие витамина D (см. рисунки 2 (e) и 2 (f)).Мы наблюдаем, что при очень высоких значениях krese или очень высокой патогенной стимуляции (kinp) система всегда оказывается в слабо регулируемом состоянии, когда эффекторные Т-клетки в изобилии, даже при наличии стандартного уровня витамина D. Однако витамин D помогает сохранить требуемую (умеренную / бистабильную) регуляцию в большом диапазоне krese и действительно контролирует широкий диапазон патогенной силы (kinp). В зависимости от интенсивности патогенной стимуляции и роста эффекторных Т-клеток, опосредованного активацией APC, иммунная система устанавливает сбалансированную регуляцию для контроля воспаления.Этот результат неизбежно предполагает важную роль витамина D в включении такой необходимой регуляции.
В свете предыдущих результатов стоит упомянуть здесь, что бистабильность является ключевой концепцией для понимания основных явлений клеточного функционирования [42,43]. Интересно, что в присутствии витамина D бистабильность становится более устойчивой к значительным изменениям патогенной стимуляции. С помощью классификации трех областей регулирования (слабая, умеренная, сильная) мы исследовали границы между любыми двумя.Как и на предыдущем графике, здесь мы одновременно варьировали как скорость патогенной стимуляции (kinp), так и скорость активации APC, опосредованной эффекторными Т-клетками (krese) (см. Рисунок 3). Необходимо отметить, что здесь образование активных АПК играет центральную роль в определении площади бистабильной области. В отсутствие витамина D продукция активных APC регулируется слабым ингибирующим действием регуляторных Т-клеток. Таким образом, усиленное производство активных APC особенно ответственно за появление такой ограниченной бистабильной области (рис. 3 (а)).В присутствии витамина D активный
Рис. 2. Вариация концентрации Т-клеток при регулировании от слабого до сильного. Установившиеся концентрации эффекторных Т-клеток (TEff, показаны красным) и регуляторных Т-клеток (TReg, показаны зеленым) нанесены на график в зависимости от различных диапазонов патогенной стимуляции (kinp) в трех различных регуляторах APC-опосредованных эффекторных Т-клеток ( Кресе). Мы находим стабильное строго регулируемое состояние при кресе ~ 10 как (а) в отсутствие витамина-D, и (б) в присутствии витамина-D.Строгая регуляция остается сильной даже в присутствии витамина D при том же значении кресе. Бистабильное состояние появляется при krese ~ 102, где как слабо регулируемое состояние (показано пунктирной линией), так и строго регулируемое состояние (показано сплошной линией) могут сосуществовать (c) в отсутствие витамина-D и (d) в присутствии витамина -D. Стабильное слабо регулируемое состояние появляется при кресе ~ 103 (е) в отсутствие витамина D. (f) В присутствии витамина D бистабильное состояние расширяется в широком диапазоне предельного значения крезе. За этим пределом он попадает в слабо регулируемый режим.Обратите внимание, что здесь мы рассматриваем константы скорости, связанные с витамином D, как kaDt ~ 10 7, keDt ~ 10 3 и другие значения скорости, приведенные в таблице 1. Оптимальная концентрация витамина D означает стабильное значение витамина D (-50 нмоль / л).
Популяция
APC значительно уменьшилась из-за комбинированного воздействия регулирующих T-клеток и витамина D. Рисунок 3 (b) дает четкое описание того, что в присутствии стандартного (оптимального) уровня витамина D бистабильная область расширяется из-за снижения скорости процесса активации APC, опосредованного эффекторными Т-клетками.Однако из рисунка очевидно, что в присутствии витамина D слабая регуляция смещается в сторону больших значений скорости активации APC, опосредованной эффекторными Т-клетками. Результат свидетельствует о силе витамина D, который не позволяет иммунной системе выйти за пределы взрывоопасного предела активности эффекторных Т-клеток.
Изменение иммунологических компонентов во времени
В отсутствие витамина D. Мы наблюдали некоторые интересные результаты при изучении анализа временной эволюции иммунологических компонентов в вышеупомянутых трех областях регуляции.
Рис. 3. Влияние витамина D на фазовую диаграмму иммунной регуляции. (a) В отсутствие и в присутствии витамина-D пара скоростей регуляции, (i) скорость активации APC, опосредованная патогенной стимуляцией (kinp), и (ii) скорость активации APC, опосредованная эффекторными Т-клетками (krese ) варьируются, чтобы определить границы между тремя конкретными областями регулирования: слабой, умеренной и сильной. (а) В отсутствие витамина D интервалы слабой регуляции занимают гораздо более широкую область, в то время как интервалы умеренной (бистабильное состояние) и сильной регуляции относительно малы.(б) В присутствии витамина D границы фаз смещаются: сильные области становятся шире. Бистабильные регионы относительно расширяются. Слабые области становятся значительно сжатыми, чем это происходит в отсутствие витамина D. Рассмотренные здесь константы скорости аналогичны рисунку 2.
Здесь мы представили динамические изменения элементов в зависимости от времени (дней), которые количественно объясняют некоторые признаки иммунных ответов. В отсутствие витамина D (рис. 4 (а)) в течение нескольких часов мы видим резкое увеличение количества эффекторных Т-клеток, достигающее пикового значения.Как было сказано ранее, это обычно относится к началу адаптивного иммунного ответа. Это согласуется с большинством экспериментальных результатов, которые предполагают, что распознавание и, следовательно, активация / начало адаптивного ответа происходит в течение нескольких часов после патогенного вторжения [44,45].
Патогенный рост начинает умирать гораздо быстрее сразу после начала выработки эффекторных Т-клеток. Теперь у нас есть огромная популяция эффекторных Т-клеток, которые были активированы из наивных Т-клеток после активации APC.Популяция этих Т-клеток остается значительно выше даже после того, как нагрузка патогенов становится значительно подавленной. Таким образом, нерегулируемый взрыв производства эффекторных Т-клеток часто вызывает различные виды аутоиммунных заболеваний [13-15,41,46].
Рисунок 4. Временная эволюция иммунного ответа. Динамическое изменение патогенов и эффекторных Т-клеток рассчитывается как (а) в отсутствие, так и (б) в присутствии оптимального уровня витамина D. В обоих случаях адаптивный ответ наступает через несколько часов после патогенного вторжения.Процесс аннигиляции патогена начинается после распознавания патогена APC и последующей APC-опосредованной активации Т-клеток. (а) В отсутствие витамина D только слабый, ингибирующий контроль регуляторных Т-клеток на продукцию эффекторных Т-клеток приводит к повышенной постоянной концентрации TEff-клеток. Это может увеличить риск аутоиммунных заболеваний. (б) Присутствие витамина D оказывает больший контроль над производством клеток TEff. После устранения патогенной нагрузки количество эффекторных Т-клеток также значительно снижается.Это может снизить риск аутоиммунных заболеваний. В то же время обратите внимание на возможность повторного вхождения патогена, который до определенного уровня способствует формированию адаптивной толерантности иммунной системы. (В обоих случаях kinp, krese выбраны так, чтобы они оставались в бистабильной области (kin, = 10, krese = 100), как показано на рисунке 3. Другие рассматриваемые здесь константы скорости аналогичны рисунку 2.
После устранения патогенной нагрузки через долгое время (около 10-20 дней) развивается новое устойчивое состояние.Как только нагрузка патогена снижается, организм создает иммунологическую память об этом конкретном патогене, что соответствует постоянному значению эффекторных Т-клеток. Это может быть особенно полезно, если тот же патоген поразит снова. Тогда иммунный ответ будет быстрым и более эффективным в подавлении целевых патогенов [47]. Результат также означает, что в отсутствие витамина D постоянное значение эффекторных Т-клеток приближается к пределу, при котором существует высокий риск развития аутоиммунного расстройства.
В наличии стандартный уровень витамина D. Витамин-D играет решающую роль в возникновении адаптивного ответа. Он изменяет сценарий, как описано в последнем подразделе. Как только популяция Т-эффекторов начинает увеличиваться, производство активного витамина D [D *] усиливается. Это, в свою очередь, активирует рост регуляторных Т-клеток, которые вместе с [D *] регулируют агрессивные воспалительные реакции, проявляемые Т-эффекторными клетками, восстанавливая контроль в организме. В этом процессе популяция эффекторных Т-клеток расслабляется гораздо быстрее (рис. 4 (б)).В результате значительно снижается уровень уничтожения патогенов. В нашей модели активации витамина D
начинает быстро расти в течение 1 или 2 дней. Таким образом, мы обнаружили, что активный витамин D не играет существенной роли на самой начальной стадии роста или распада патогенов. В присутствии витамина D мы наблюдаем возможность повторного входа патогена, который может сохраняться в течение длительного времени [12].
Чтобы компенсировать между эффективным очищением от нагрузки патогенов и риском аутоиммунных заболеваний, витамин D играет роль отрицательного катализатора в производстве эффекторных Т-клеток.Как следствие, в присутствии витамина D скорость аннигиляции патогенов в течение более длительного времени также снижается. Следовательно, из наших анализов мы обнаруживаем, что при оптимальном регулировании витамина D, чтобы минимизировать риск аутоиммунных заболеваний, наша иммунная система обязана терпеть некоторое количество патогенов. Фактически, здоровая иммунная система всегда характеризуется устойчивостью к определенной степени патогенной стимуляции. Тот факт, что витамин D был задействован как важный фактор в нескольких различных аутоиммунных заболеваниях, сохраняя бистабильность, предполагает его эффективность в контроле самотолерантности организма [48-50].Здесь стоит упомянуть, что экспериментальные наблюдения, связанные с адаптивной передачей толерантности, также подтверждают появление такой би-стабильности, при которой в системе сохраняется сбалансированное сосуществование сильных и слабо регулируемых иммунных ответов [51].
Анализ устойчивого состояния и оптимальный витамин D
Одна важная деталь, которую необходимо здесь учитывать, — это появление нового устойчивого состояния в присутствии витамина D с его строго контролируемым гомеостазом.Чтобы понять важность витамина D в вышеуказанном ответе, были рассмотрены различные начальные концентрации витамина D, [Din0]. Таким образом, мы рассмотрели различные начальные концентрации витамина D от 10 4 до 10 нмоль / л. Было получено изменение уровней Т-клеток и уровней патогенов в недавно установленном устойчивом состоянии, и эти концентрации нанесены на график в зависимости от log [Din0] на рисунке 5. Для измерения оптимального диапазона витамина D нам также необходимо контролировать иммунную регуляцию. как патогенная устойчивость, поскольку они тесно связаны.Важно отметить, что мы не можем установить такую строгую регуляцию витамином D, за которой иммунная система развивает большую патогенную толерантность, а клиренс патогенов эффекторными Т-клетками незаметно не удается.
Эффекты локального превращения неактивного 25 (OH) D3 в активный 1,25 (OH) 2D3, опосредованного DC, на последующие Т-клеточные ответы были измерены проточной цитометрией, и результаты были подробно проанализированы Jeffery et al. [52]. Они изучили, как это преобразование может способствовать развитию противовоспалительного Т-клеточного фенотипа (такого как CTLA-4) и подавлять воспалительную экспрессию IL-17, IFN-y и IL-21.Дозозависимые вариации таких Т-клеточных ответов показаны на рис. 2. (F) в упомянутой статье [52]. Тенденция ответных изменений вместе с концентрацией 25 (OH) D3 довольно хорошо согласуется с результатами, изображенными на рисунке 5, которые мы получаем из расчета нашей модели. Следуя их подсказке, в настоящем исследовании мы также рассматриваем циркулирующую неактивную форму витамина D (25 (OH) D3) как эффективный маркер статуса витамина D. Наши зависимые от дозы кривые также совпадают с экспериментальными данными Correale et al.[25].
Для приведенного выше набора данных мы обнаружили, что оптимальный уровень витамина D находится в диапазоне 50-100 нмоль / литр , где уровни патогенных и эффекторных Т-клеток остаются в диапазоне достаточно низкого риска. Недавно большое количество эпидемиологических исследований и комитет Института медицины США сообщили, что уровень 25-гидроксивитамина-D в сыворотке> 20 нг / мл (50 нмоль / л) является желательным для костей и общего состояния здоровья [53–55]. Эти исследования рекомендуют как верхний, так и нижний пределы безопасного потребления витамина D.Высокие уровни IgE наблюдались при очень низком уровне 25-гидроксивитамина-D3 (<10 нг / мл или <25 нмоль / л).
Рис. 5. Анализ значений устойчивого состояния как функция логарифма (начальный уровень витамина D). Оценка устойчивого состояния регуляции с точки зрения концентраций патогена [P], эффекторных Т-клеток [TEff] и регуляторных Т-клеток [TReg] при различном начальном потреблении витамина D. В районе значения концентрации витамина D 50 нмоль / л концентрация TEff падает ниже значения концентрации TReg, чтобы установить строгую регуляцию, необходимую для предотвращения аутоиммунных заболеваний.Значение установившегося состояния патогенов начинает увеличиваться, даже если значение TEff превышает [Din0] = 100 нмоль / л. Мы указываем (серой лицевой полосой) оптимальный диапазон витамина D от 50 до 100 нмоль / литр, при котором уровни патогенных и эффекторных Т-клеток остаются на достаточно низком уровне. Уровень витамина D выше 100 нмоль / литр соответствует тревожной концентрации по сравнению со стандартным пределом витамина D. и при очень высоких уровнях 25-гидроксивитамина-D3 (> 135 нмоль / л) [54].
Другое важное исследование показало, что высокая концентрация 25 (OH) D3 (более или = 100 нмоль / л) часто приводит к статистически значимому (2-кратному) увеличению риска рака поджелудочной железы [55,56].Таким образом, настоящее исследование дает оценку в правильном диапазоне оптимальной концентрации витамина D.
Чувствительность к набору параметров, связанных с витамином D
Чтобы исследовать устойчивость и чувствительность констант скорости, связанных с витамином D, необходимо тщательно изучить их эффекты в широком диапазоне. Дополнительная причина для подтверждения чувствительности заключается в том, что эти значения варьируются от системы к системе (здесь от человека к человеку), и значения могут колебаться даже для одного и того же человека в зависимости от различных условий.Хотя точное количество констант скорости может варьироваться, эффективный тренд должен сохраняться в пределах определенного диапазона.
Поскольку как активные APC, так и эффекторные Т-клетки модулируются воздействием активного витамина D, мы исследовали результаты различных возможностей комбинации kaD ™ и keD * (определенные в таблице 1). Из рисунка 5 очевидно, что для получения безопасной границы воздействия витамина D нам необходимо эффективно проверять как концентрацию эффекторных Т-клеток, так и концентрацию патогенов.Здесь мы просканировали пространство параметров, чтобы различить разные зоны в зависимости от популяции патогенных и эффекторных Т-клеток. Однако при высоких концентрациях витамина D рост патогенов может усиливаться из-за подавления продукции эффекторных Т-клеток. Здесь пространство параметров (logkaDt, logkeDt) предполагает, что профиль патогенных и эффекторных Т-клеток менее чувствителен к keD *. Он скорее показывает значительную вариацию с изменением kaD „. Этот анализ показывает две отдельные области: (i) В области низкого воздействия витамина D (kaD »* 10-8-10-4) мы получаем зону поражения патогенов, где концентрация патогенов незначительна, но в то же время в диапазоне kaD * * 10-8-10-7 существует зона обострения эффекторных Т-клеток.(ii) В области очень сильного воздействия витамина D (kaD * * 10-1-102) мы получаем зону поражения эффекторных Т-клеток. Здесь мы находим зону рецидива патогена, где стабильная концентрация патогена остается значительно большей, когда система гиперрегулируется витамином-D. Диапазон между kaD, * 10-7-10-4, а также keD, * 10-6-10-2 — это оптимальное пространство параметров для активного воздействия витамина D, позволяющего избежать роста патогенных и эффекторных Т-клеток (см. Рис. 6). ).
Обсуждение и сводка результатов
Недавние экспериментальные исследования предоставили большое количество количественной информации о иммуомодулирующих функциях витамина D и установили эти функции, выходящие за рамки его четко заявленной роли в метаболизме кальция [19-25].Чтобы понять эти недавние эксперименты, мы разработали теоретическую крупнозернистую модель, основанную на этой сети взаимодействия. Сеть динамически соединяет различные иммунные компоненты, которые, как было экспериментально установлено, участвуют в регулируемых витамином D иммунных ответах. Сформулированная кинетическая схема описывает эволюцию во времени этих компонентов, которые в основном включают патоген, витамин D, APC, эффекторные Т-клетки и регуляторные Т-клетки. Здесь мы суммируем соответствующие наблюдения, которые возникли из модели кинетической сети.
(i) Анализ устойчивого состояния настоящей кинетической схемы устанавливает три предела регулирования: слабый, умеренный и сильный, как в отсутствие, так и в присутствии витамина D. Фазовые диаграммы разделенных границей трех областей иммунной регуляции показывают, что в присутствии оптимального витамина D область сильной регуляции становится широкой, а область умеренной (или бистабильной) регуляции становится более протяженной. Слабая регуляторная область смещается в сторону более высоких значений скорости активации APC, опосредованной эффекторными Т-клетками (krese), и становится более ограниченной, чем та, которая наблюдается в отсутствие витамина-D.Это исследование предлагает полуколичественную картину, подтверждающую несколько экспериментальных и клинических наблюдений, которые показывают, как витамин D регулирует иммунную систему, ограничивая ее функцию в пределах от сильного до умеренного регулирования, что значительно снижает риск аутоиммунных заболеваний [11-15,46].
(ii) Анализ динамики иммунологических компонентов во времени однозначно показывает достижение нового устойчивого состояния в присутствии оптимального уровня витамина D. Динамическая характеристика задействованных компонентов показывает, что для распознавания патогенного роста требуется несколько часов, и этот факт в целом согласуется с большинством экспериментальных результатов [44,45].После активации витамина D избыточная популяция эффекторных Т-клеток расслабляется до сравнительно более низкого значения (когда мы включаем эффекты оптимального витамина D). Но такое нисходящее регулирование для предотвращения аутоиммунных заболеваний происходит за счет возможности повторного входа, в определенной степени, патогена, что снова повышает толерантность здоровой иммунной системы. Важность витамина D в контроле толерантности также подтверждена экспериментально.
(iii) Количественные предсказания настоящей модели хорошо согласуются с несколькими недавними экспериментальными исследованиями и клиническими наблюдениями [12,25,44-50,52-58].Мы попытались количественно определить, сколько витамина D необходимо, чтобы противостоять аутоиммунитету и почему? Наши дозозависимые вариации Т-клеточного ответа вместе с концентрацией витамина D, по-видимому, отлично коррелируют с экспериментальными данными Jeffery et al. и Correale et al. [52,25]. Кроме того, мы обнаружили, что безопасный диапазон витамина D в основном определяется взаимосвязью патогена, эффекторных Т-клеток и регуляторных Т-клеток. Диапазон ограничен как гиперрегуляцией, так и воспалением эффекторных Т-клеток.Совсем недавно рандомизированные контролируемые исследования (РКИ) предполагают, что следует предусмотреть элемент осторожности в отношении рекомендации высоких концентраций 25 (OH) D3 в сыворотке крови в качестве рутинной клинической практики и распространения среди всего населения [56-58]. Это говорит о том, что требуются более активные совместные усилия, а также экспериментальные и теоретические инициативы.
(iv) Регуляторное влияние активного витамина D на APC и эффекторные Т-клетки исследуется здесь с помощью анализа устойчивого состояния. Мы обнаружили, что нелинейная регуляция витамина D
Рисунок 6.Влияние активного витамина D на устойчивые профили патогена [P] и эффекторных Т-клеток [TEff]. (a) Мы изменяем одновременно влияние активного витамина -D ([D *]) на APC (kaDt) и эффекторные Т-клетки (keDt). Мы находим разные области: (i) При низком воздействии витамина D (kaD * * 10-8-10-4) мы получаем зону поражения патогена, но зону обострения TEff (kaD, * 10-8-10-7). (ii) При высоком воздействии витамина D (kaD * * 10–1-102) в устойчивом состоянии концентрация патогена значительно увеличивается, что выделяется как зона рецидива патогена.Однако в зоне рецидива патогена мы находим зону поражения TEff. В качестве основных параметров значения принимаются kin, = 10, krese = 102. Значения остальных параметров взяты из таблицы 1.
Таблица 1. Значения основных параметров (* продолжительность принята в «днях»).
чувствителен к работе APC. Этот конкретный прицельный параметр во многом определяет возникновение и диапазон бистабильности. Ранние экспериментальные исследования также сообщают о таком заметном влиянии профиля созревания и активации DC в присутствии витамина D [21].
Как мы упоминали ранее, анализ устойчивого состояния предложенных основных уравнений показывает сложные отношения между уровнями витамина D и Т-регуляторными клетками, поддерживаемыми гомеостазом. Эти отношения предполагают, что при гомеостазе более низкие уровни витамина D соответствуют более низкой популяции Т-регуляторных клеток, что снова предполагает, что, как только патоген попадает в организм, характер иммунного ответа, как ожидается, будет менее регуляторным и, следовательно, более сильным. воспалительный или агрессивный. Кроме того, в условиях слабой регуляции мы изучили временную прогрессию как регуляторных, так и эффекторных Т-клеток.Интересно, что мы обнаруживаем сопряженную колебательную динамику эффекторных Т-клеток (TEff) и регуляторных Т-клеток (TReg), которые начинают развиваться в течение 2-5 дней и периодически продолжаются. В присутствии патогена, когда система стремится к немного слабому режиму регуляции, мы наблюдаем динамическую перекрестную регуляцию во временной прогрессии популяции регуляторных и эффекторных Т-клеток. Это описано в тексте S2 в файле S1 и представлено на рисунке S1 в файле S1 [16]. Влияние внутреннего осцилляторного поведения, связанного с витамином D, на эффекторные Т-клетки может служить ярким признаком фенотипа заболевания в клинической терапии [29].
Критическую роль различных клеток, участвующих в иммунном ответе, особенно неактивных и активных концентраций витамина D, можно понять, исследуя динамику ответа. Мы действительно осознаем тот факт, что количественные результаты анализа in vivo воздействия высоких доз витамина D или любого его стероидного аналога несколько неоднозначны. Последствия смертельных заболеваний как низкой, так и очень высокой дозы витамина D относительно хорошо изучены. Мы также осознаем сохраняющуюся дилемму точного определения недостаточности витамина D и трудности с определением безопасного диапазона.Наш модельный расчет эффективно определяет, что существует тонкий интервал концентраций витамина D, который имеет решающее значение для поддержания надлежащего ответа на патоген. Чрезвычайно низкие уровни витамина D могут привести к повышенному риску аутоиммунных реакций, а чрезвычайно высокие уровни предполагают чрезвычайно толерантный ответ, который может увеличить риск опухолей и роста раковых клеток, а также различных аллергических реакций, вызванных повышенными концентрациями IgE [57, 58].
Важно отметить, что два фермента CYP27B1 и CYP24A1 и популяция VDR играют важную роль в балансировании нескольких иммунологических ответов. Дефект или недоступность любого из этих белков сильно нарушит всю иммунологическую сеть. Ряд процессов, опосредованных D * -VDR, которые имеют огромные последствия, еще полностью не изучены. Неисправность этих ферментов (таких как: CYP27B1 и CYP24A1) также может отражать более глубокую проблему (например, генетические мутации), которую трудно исправить [59,60].Это явно требует более количественного анализа.
Здесь стоит упомянуть, что активация наивной Т-клетки в эффекторную или регуляторную Т-клетку также является сложным процессом. Это начинается со сканирования поверхности APC в лимфатических узлах на предмет молекул типа MHC класса II с помощью наивных Т-клеток. Если определенный эпитоп распознается и присутствуют костимуляторные молекулы, то запускается процесс активации [61,62]. Теперь это можно понять с помощью анализа энергетического ландшафта.Процесс успешной активации можно рассматривать как преодоление Т-клеткой барьера в энергетическом ландшафте. Это может быть достигнуто либо через один успешный контакт с APC, либо за несколько контактов, если второй или последующий контакт происходит в течение конечного времени. Если Т-клетка находится выше сепаратрисы в энергетическом ландшафте, то вероятность успешной активации выше, которая присутствует только в течение конечного времени после предыдущего возбуждения. Приведенная выше картина аналогична иммунологическим исследованиям, проведенным Hong et al.[63] и Das et al. [62] и модель ферментного катализа, предложенная ранее Мин, Се и Багчи [64]. Однако, чтобы сделать настоящую модель управляемой, нам пришлось игнорировать такую сложность активации Т-клеток.
Используемый здесь подход главного уравнения был решен как с помощью детерминированного, так и стохастического подхода с учетом начальных значений параметров и потоков. Внутри биологической клетки всегда могут быть большие колебания из-за факторов окружающей среды или других причин [65,66]. Такие колебания могут вызвать переход от слабого регулирования к сильному.Это вопрос, заслуживающий дальнейшего изучения.
Хотя наша модель является крупнозернистой, а полученные результаты являются полуколичественными из-за отсутствия некоторых кинетических параметров, это исследование, возможно, представляет собой первое теоретическое исследование роли витамина D в иммунной регуляции. Несмотря на его ограничения, мы считаем, что кинетическое взаимодействие между патогеном, эффекторными Т-клетками и неизбежное участие витамина D останутся основными ингредиентами в предстоящих исследованиях.
В будущем мы планируем расширить нашу систему уравнений, включив в нее эффекты лекарств, таких как иммунодепрессанты (например,g., глюкокортикоиды), которые вносят дополнительную конкуренцию в реакционную сеть [13].
Методы
Разработка модели крупнозернистой реакционной сети
Чтобы описать сложное взаимодействие между различными типами иммунных клеток, патогенами и модулирующей ролью витамина D, сначала нам нужно разработать простой грубый подход, который можно решить и понять. Сложность возникает из-за большого количества биохимических механизмов в организме человека, которые сильно связаны друг с другом [67,68].Понимание взаимосвязи между этими различными механизмами, включающими разные типы клеток, может в конечном итоге потребовать детализации на молекулярном уровне. Здесь предлагается более простая, хотя и более грубая версия, которая объясняет некоторые из сложностей, присутствующих на молекулярном уровне, путем их крупномасштабного анализа на клеточном уровне. Наглядное описание исходной сложной сети и связанной с ней крупнозернистой сети показано на рисунке S2 в файле S1 и рисунке S3 в файле S1 соответственно [16].С этой целью мы проводим анализ моделей на основе активации, деактивации и регуляции Т-клеток, следуя некоторым экспериментальным результатам, обсуждаемым ниже.
- (i) Миелоидные дендритные клетки (также мы называем их антигенпредставляющими клетками (APC)), присутствующие в различных органах, являются ключевыми игроками, участвующими в запуске адаптивного иммунного ответа. После созревания и презентации патогена эти дендритные клетки служат для активации наивных Т-клеток в эффекторные Т-клетки. Напротив, незрелые дендритные клетки при контакте с патогеном превращают наивные Т-клетки в регуляторные Т-клетки в отсутствие сигнала созревания [19,20].
- (ii) Эффекторные Т-клетки выделяют цитокины, которые повышают активность фермента 1 α-гидроксилазы (CYP27B1), который, в свою очередь, индуцирует превращение активного витамина D из его неактивной формы. Здесь стоит упомянуть обширное экспериментальное исследование Correale et al. которые сообщили, что CD4 + Т-клетки способны метаболизировать 25 (ОН) -витамин D до 1,25 (ОН) 2-витамина D, что снова подавляет функцию Т-клеток [25].
- (iii) Активная форма витамина D, 1, 25 (OH) 2D3 [D *] модулирует иммунный ответ посредством ингибирования дифференцировки DC и созревания в мощные APC [21].
- (iv) Повышенная продукция [D *], непосредственно подавляет продукцию эффекторных Т-клеток [TEff] и усиливает ответ регуляторных Т-клеток [TReg] CD4 + / CD25 + / FoxP3 +. Эти клетки TReg также эффективно ингибируют пролиферацию клеток TEff [22-25].
Грубая детализация сети взаимодействия достигается за счет нескольких упрощающих наблюдений и важных предположений. Вот они:
(a) Клетки Th2, Th3 и Th27 сгруппированы вместе как эффекторные Т-клетки. Подробное описание этих Т-клеток представлено в тексте S1 в файле S1 [16].
(b) Хорошо известно, что первичное молекулярное действие 1,25 (OH) 2D3 заключается в инициации транскрипции гена путем связывания с VDR, который является членом суперсемейства рецепторов стероидных гормонов факторов транскрипции, активируемых лигандом. Таким образом, VDR является важным фактором функций, опосредованных 1,25 (OH) 2 D3. Более подробную информацию о VDR можно найти в ссылке 69 [69].
Напротив, есть сообщения, что 1,25 (OH) 2D3 также обладает быстрыми действиями, которые по существу не опосредуются посредством транскрипционных событий с участием VDR.Фактически это действия, инициируемые мембраной [70]. В данной модели мы не учли эффект VDR. Мы рассмотрели только производство активного витамина D из его неактивной формы при активации Т-клеток.
(a) В обращении неактивная форма витамина D, 25 (OH) D3, обычно используется в качестве индикатора статуса витамина D. Однако в дендритных клетках (ДК) использование этого предшественника зависит от его поглощения клетками и последующего превращения ферментом CYP27B1 в активный [D *] [71]. [D *] имеет жесткий контроль над скоростью производства гомеостаза, который саморегулирует его производство, напрямую регулируя активность цитохрома P450 CYP24A1.В нашей модели мы рассмотрели уровень неактивного витамина D в устойчивом состоянии, полученный в результате экспериментальных и клинических измерений, при этом сохраняя концентрацию этих ферментов в качестве неявных факторов.
В данном контексте мы рассматриваем следующий набор биологических преобразований. Большинство из них являются каталитическими реакциями с точки зрения повышающей или понижающей регуляции.
(1) Первичный шаг — уничтожение патогена эффекторными Т-клетками.
Патоген (P) + эффекторная Т-клетка (TEff)? Pkiiied + TEff (i)
(2) Производство эффекторных Т-клеток требует присутствия активной антигенпрезентирующей клетки (APC).Активный APC, с другой стороны, продуцируется следующей последовательностью реакций. 1-й АРС в состоянии покоя образуется в результате взаимодействия неактивных АРС и патогена.
Неактивный APC (Ain) Z P? Остающийся APC (ARes) Z P (ii)
AAct Z TReg? ARes Z TReg AActZD *? AResZD *
(3) Дальнейший контакт с патогенами и / или контакт с эффекторными Т-клетками способствует превращению покоящихся АРС в активные АРС.
ARes ZP? AActZP ARes Z TEff? AAct Z TEff
(4) Затем эффекторная Т-клетка продуцируется взаимодействием между предшественником / наивной Т-клеткой с активным APC.
Наивные Т-клетки (TNa) Z Активные APC (AAct)? TEff Z AAct (iv)
(5) Одновременно неактивный витамин D превращается в активный витамин D при контакте с эффекторными Т-клетками.
Неактивный витамин — D (Din) ZTEff -Активный витамин — D (D *) ZTEff (v)
Основные уравнения для количественной оценки динамики реакционной сети
А теперь сделаем еще несколько важных предположений, прежде чем мы приступим к написанию основных уравнений:
(i) Для Патогена, неактивных APC и наивных Т-клеток, каждый имеет коэффициент рождаемости, который включает в себя уровни притока и пролиферации, а также коэффициент смертности, аналогичный уровню распада, который включает естественную гибель клеток.Уровень смертности каждого компонента линейно зависит от его концентрации.
(ii) Предполагается, что все вероятности перехода постоянны во времени, но могут варьироваться от системы к системе (т.е. здесь от человека к человеку) в зависимости от применяемых условий.
(iii) Чтобы масштабировать единицу, здесь мы предполагаем, что в отсутствие патогена сотня (среднее количество Т-клеток, присутствующих в сотне нанолитровых образцов крови) предшественников Т-клеток уже существует.
Вышеупомянутые аннигиляция, рекомбинация и каталитические реакции приводят к следующему набору связанных основных уравнений.Уравнения обширны. Фактически, возможность увеличения размера является критически важным фактором устойчивости нашей модели.
dP dt = sP — mPP — кП TEffP
(1)
(6) Отдыхающие Т-клетки и витамин D могут инициировать образование регуляторных Т-клеток из наивных Т-клеток.
dAin dt = SA kinpAinP maAin
(2) Наивные Т-клетки (TNa) ZResting APC (ARes) — TRegZARes
\ Наивные Т-клетки (TNa) ZActive Viatmin-D (D *)? TRegZD =
(7) Как регуляторные Т-клетки, так и активный витамин D могут подавлять выработку эффекторных Т-клеток, чтобы контролировать гиперактивность иммунной системы.
дт
= kinpAinPZ kay pRegAAct Z kaD * AActD
(3)
— КРЕСЕАРЕС ТЭФ — КИНПАРЕС — МААРЕС
[t i t. т убил я т ~
TEffZ TReg- TEff Z TReg 1 TEff ZD *? TEffkilledZD *
дт
= kyeseARes TEff Z knpAResP — kay pRegAAct
(4) — kaD * AActD * —maAj
(8) Цикл завершается преобразованием активного APC в состояние покоя снова той же парой, TReg и D *, которые работают на
тандем.
= Sp —kanAActTNa — kyesnAResTNa — knD * TNaD * —mnTffa (5)
dTEff
дт
= kanAAct TNa — ker TEff TReg — keD * TEffD * — me TEff
(6) dTReg
дт
= kresnARes TNa Z knD * TNaD * — mr TReg
(7) dt = sD keDTEffDin mDDin
(8) дД
= keD TEff Din — mD * D =
(9)
Где термины означают следующее:
kij R Коэффициенты вероятности перехода,
Sk R Производительность по корпусу компонента k,
mi R Общая смертность по компоненту i,
P R Концентрация патогена,
Ain R Концентрация неактивных антигенпрезентирующих клеток без захвата патогена,
ARes R Концентрация покоящихся антигенпрезентирующих клеток после захвата патогена,
AAct R Концентрация активированных антигенпрезентирующих клеток после распознавания патогена и контакта с эффекторными Т-клетками.
TNa R Концентрация наивных Т-клеток,
TEff R Концентрация эффекторных Т-клеток,
TReg R Концентрация регуляторных Т-клеток,
Din R Концентрация неактивной формы витамина D3 (1,25 (OH) D) в организме.
D * R Концентрация активной формы витамина D3 (1,25 (OH) 2D) в организме
То есть мы использовали одну и ту же букву для обозначения и вида, и его концентрации. Это не должно вызывать недоумения.
Системные параметры и анализ данных
Систему из девяти связанных дифференциальных уравнений сложно решить аналитически.Мы получаем зависящие от времени концентрации всех компонентов, участвующих в схеме, используя хорошо известный стохастический симуляционный анализ, предложенный Гиллеспи [[38]. С помощью этого метода были изучены как одиночный молекулярный, так и ансамблевой ферментный катализ. Все результаты, представленные в этой статье, получены с использованием метода стохастического моделирования. Однако мы также проверили согласованность каждого результата, используя детерминированный подход, который легче реализовать.
Здесь мы рассмотрели 100 нанолитров образца крови.В отсутствие патогена этот образец крови эффективно содержит стационарную концентрацию всех клеток-предшественников [72-74]. Поскольку все реакции являются бимолекулярными, ожидается, что проблема будет зависеть от объема реакции. Таким образом, мы сохранили фиксированный объем ящика на уровне ста нанолитров, а все константы скорости указаны в единицах дня. Мы внимательно следили за формализмом, развитым в [3]. 30.
Кроме того, мы предположили, что в отсутствие антигена сотни Т-клеток-предшественников могут существовать в этом фиксированном объеме (100 нанолитров) в соответствии с известными экспериментальными значениями [72,73].Эти Т-клетки имеют оборот 1% в день. Также нормализуется концентрация возбудителей и АПК. Скорость производства и смертность этих компонентов задаются таким образом, чтобы их значения в стационарном состоянии стали едиными. Другие связанные вероятности / константы скорости различных наборов реакций используются из ранних работ в этой области [30]. Однако для витамина D константы продуцирования и смертности рассчитываются исходя из их стационарной концентрации. Другие константы скорости, связанные с витамином D, рассматриваются в нашем исследовании как переменные, поскольку у нас нет экспериментальных данных о них.Реально такая модель требует оценки нескольких значений параметров скорости. К сожалению, очень трудно определить точные значения некоторых из этих констант скорости. Такие параметры скорости зависят от нескольких факторов и различаются от вида к виду. Таким образом, у них нет определенного стандартного значения. Например, было бы довольно сложно определить уровень смертности эффекторных и регуляторных Т-клеток, поскольку в настоящей модели эти параметры скорости также включают скорость пролиферации вместе с уровнем их смертности.Более того, патогенная стимуляция может быть различной в зависимости от их силы и характера.
Следовательно, основная трудность прогнозных теоретических исследований в этой области заключается в отсутствии точных значений констант скорости / вероятностей перехода. В настоящем исследовании мы использовали следующий подход, чтобы обойти эту трудность. (i) В некоторых случаях, когда значения можно было оценить из литературы, мы использовали известное значение и варьировали его в диапазоне, чтобы проверить чувствительность результатов.(ii) В некоторых случаях использовались оценки по порядку величины [30]. Мы также сосредоточились на изучении фазовой диаграммы, варьируя некоторые ключевые параметры скорости, которые не известны, и искали оптимальную область, где результаты чувствительны к пространству параметров (с учетом экспериментальных и предполагаемых значений констант скорости и концентраций). С этой целью мы варьировали константы скорости в очень широком диапазоне. Кроме того, была нормализована концентрация элементов-прекурсоров, чтобы отразить многократное изменение уровня производства.Принимая типичные значения, указанные ниже (см. Таблицу 1), в настоящей работе выполняются временная оценка системы и другие анализы. Здесь мы использовали стандартное определение устойчивого состояния, т.е. когда концентрация различных видов неизменна во времени (dc / dt = 0). В частности, для стохастического моделирования предполагается, что устойчивое состояние достигается, когда концентрация вида колеблется около среднего значения без какого-либо заметного дрейфа в течение длительного времени.
Дополнительная информация
Файл S1 содержит текст S1, который описывает процесс активации Т-клеток и введение эффекторных и регуляторных Т-клеток, текст S2, который описывает зависящее от времени колебательное поведение антигенспецифических эффекторных (TEff) и регуляторных (TReg) Т-клеток, рисунок S1, который показывает влияние витамина D на профиль эффекторных и регуляторных Т-клеток в присутствии патогена, Рисунок S2, который показывает сложное представление адаптивного иммунного ответа, и Рисунок S3, который показывает крупнозернистую сеть из Рисунка S2.
Благодарности
Мне приятно поблагодарить профессора Анджали А. Каранде и г-на Арка Бакси за многие полезные обсуждения и предложения. Мы также благодарим доктора Манту Сантру, доктора Бимана Яну, господина Кушала Багчи, госпожу Гаури Ранадив и госпожу Прити Гараи за их постоянную поддержку в этом проекте.
Авторские взносы
Задуманы и спланированы эксперименты: SR.
