
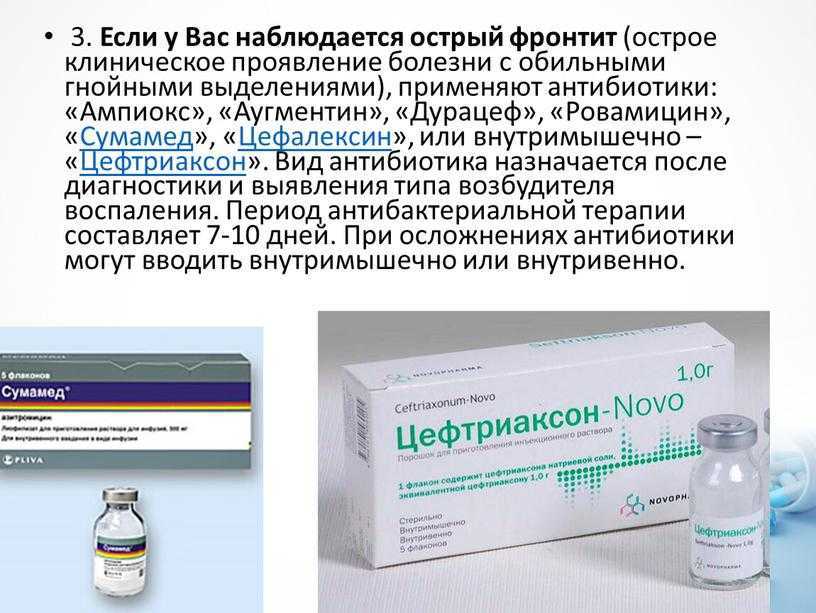
Берегите бабушек и дедушек!
содержание видео
Рейтинг: 4.0; Голоса: 1
В этом видео доктор Комаровский объяснит:
почему пожилых людей не просто желательно, а необходимо вакцинировать от детских инфекций;
какие болезни у бабушек и дедушек можно предотвратить с помощью прививок;
насколько важно, чтобы дети были вакцинированы по календарю, если они общаются со старшими родственниками
Дата: 2020-11-12
← Повышение температуры после вакцинации
Нужен ли в доме очень точный термометр? →
Похожие видео
Что такое ДЕЦИБЕЛ #5. Звук. Громкость звука. Давление звука. Сила звука.
• ElectronicsClub
Почему нужно первым заходить в самолет / Где я встречаюсь со своей старшей дочерью / Отели
• Вероника Степанова
Зависимость от капель в нос. Как избавиться?
• Доктор Комаровский
Как перестать быть славным парнем? Как вести себя с девушкой чтобы она Видела в тебе мужчину
Вот какой БЕЛОКАМЕННЫЙ город ЗАКОПАН под ПЕТЕРБУРГОМ
• Альтернативная история
ЗЕМЛЯ — глобальный КАРЬЕР прошлой ВЫСОКОРАЗВИТОЙ цивилизации
• Альтернативная история
Комментарии и отзывы: 10
MariNa
ПРОЧТИТЕ ПОЖАЛУЙСТА! Евгений Олегович, Вы говорите, пишите не стесняйтесь, я или мои помощники ответят. Вот у меня вопрос, мучающий меня, ребенку 2, 6 года, после пребывания на местом озере летом в августе переболели, как нам сказали ангиной, (высокая температура до 39, без кашля и других симптомов болезни) в итоге после лечения, назначенного педиатром попали в больницу с бронхитом, как показал снимок рентген) антибиотики кололи 7 дней. выписали домой, и спустя 2, 3 дня у ребенка температура 38, 5 37, 37, 5, 38. обратились к педиатру, выписали лечение, без антибиотиков, в течении 3-5 дней не помогало, по скорой попали снова в больницу с темп 38, 7. кололи антибиотик цифотоксим 10 дней, лежали в больнице строго. выписались. но, спустя несколько дней опять кашель сильные приступы ночью, нос с хрюкающими соплями где то внутри, и температура 37, 37, 5 постоянно держится. сдали анализы мазок из зева и носа, кровь, моча, выявили синегнойная палочка 104. назначили клацид на 10 дней и нос промывать. курс прошли но. темп все время 37 держится. нос все так же хрюкает. очень переживаю с чем это связано.
Вот у меня вопрос, мучающий меня, ребенку 2, 6 года, после пребывания на местом озере летом в августе переболели, как нам сказали ангиной, (высокая температура до 39, без кашля и других симптомов болезни) в итоге после лечения, назначенного педиатром попали в больницу с бронхитом, как показал снимок рентген) антибиотики кололи 7 дней. выписали домой, и спустя 2, 3 дня у ребенка температура 38, 5 37, 37, 5, 38. обратились к педиатру, выписали лечение, без антибиотиков, в течении 3-5 дней не помогало, по скорой попали снова в больницу с темп 38, 7. кололи антибиотик цифотоксим 10 дней, лежали в больнице строго. выписались. но, спустя несколько дней опять кашель сильные приступы ночью, нос с хрюкающими соплями где то внутри, и температура 37, 37, 5 постоянно держится. сдали анализы мазок из зева и носа, кровь, моча, выявили синегнойная палочка 104. назначили клацид на 10 дней и нос промывать. курс прошли но. темп все время 37 держится. нос все так же хрюкает. очень переживаю с чем это связано.
Yana
Спасибо Вам за ваши труды! Как же жалко, что Вас не слышит наше правительство( и мамы ярые противники вакцинации. Что самое смешное с одной такой мамой я общалась, у нас дети и собаки, меня удивило то, что собаке она делает прививки со словами если не привить собаку, то собака может умереть а ребенка нет, не прививаем, это же опасно! ) и да вет. клиники звонят и напоминают о прививках, а педиатры или детские поликлиники нет) К слову у нас мед. страховка и мы к счастью не имеем дело с гос. поликлиникой, лечение орви и тому подобное разительно отличается от лечения в гос. поликлиниках, ребенку 9лет и антибиотик мы принимали 2раза в жизни и то по результату анализов ) а детки у моих подруг которые в гос. поликлинике к сожалению часто на антибиотиках, мамы говорят педиатр так сказала, вот мы и даем а то вдруг воспаление будет или осложнение какие! Печалька
Наталі
Здравствуйте, Евгений Олегович! Просмотрела все Ваши передачи. Один вопрос у меня все-таки есть. Дочери два года. прививка от кори есть. В феврале нам надо делать очередную АКДС. У нас сместился календарь прививок, поскольку мы начали их делать после первого года жизни. Мы проживаем в Закарпатье. У нас закрыли школы на карантин из-за кори. Вопрос вот в чем: мы прививку делаем не в местной амбулатории, а в самом Ужгороде. Наша врач побуждает нас делать прививку именно сейчас. Но мне страшно ехать в автобусе три часа в одну сторону, когда у нас бушует корь. Что Вы посоветуете: можно смело ехать или стоит переждать, пока не снимут карантин с садиков и школ? Второе: хочу поблагодарить Вас за информацию о том, что и взрослым надо делать прививки от дифтерии. Мужа привили, себе маме сделаем в следующем месяце. Спасибо Вам! Здоровья Вам!
Один вопрос у меня все-таки есть. Дочери два года. прививка от кори есть. В феврале нам надо делать очередную АКДС. У нас сместился календарь прививок, поскольку мы начали их делать после первого года жизни. Мы проживаем в Закарпатье. У нас закрыли школы на карантин из-за кори. Вопрос вот в чем: мы прививку делаем не в местной амбулатории, а в самом Ужгороде. Наша врач побуждает нас делать прививку именно сейчас. Но мне страшно ехать в автобусе три часа в одну сторону, когда у нас бушует корь. Что Вы посоветуете: можно смело ехать или стоит переждать, пока не снимут карантин с садиков и школ? Второе: хочу поблагодарить Вас за информацию о том, что и взрослым надо делать прививки от дифтерии. Мужа привили, себе маме сделаем в следующем месяце. Спасибо Вам! Здоровья Вам!
Nadejda
Уважаемый Евгений Олегович, очень хочу попросить помощи у вас. Моя бабушка заболела. Я даже не помню когда у нее последний раз была температура, а сейчас уже 4ый день ходит с температурой 37 — 38 с хвостиком, кашляет, постоянно чихает, насморк есть. Мы ее пытаемся вылечить так сказать народными средствами: чай с малиной, лимоном и медом, гриппофлю, горло полощет фурацилином, нос при золоженности ринорусом капает. Но ей ни чуть не лучше. К врачу идти отказывается. А я уже не знаю как ей помочь. Ей 66 лет, у нее вообще постоянные головные боли, проблемы с сосудами есть. Я конечно понимаю, что по фотографии не лечат, но может хоть что-то посоветуете? Пожалуйста. Заранее большое спасибо.
Мы ее пытаемся вылечить так сказать народными средствами: чай с малиной, лимоном и медом, гриппофлю, горло полощет фурацилином, нос при золоженности ринорусом капает. Но ей ни чуть не лучше. К врачу идти отказывается. А я уже не знаю как ей помочь. Ей 66 лет, у нее вообще постоянные головные боли, проблемы с сосудами есть. Я конечно понимаю, что по фотографии не лечат, но может хоть что-то посоветуете? Пожалуйста. Заранее большое спасибо.
Наталья
Здравствуйте Доктор. мои свекровь 67 лет и свекр 69 решили делать прививку АКДС(я им доказала что надо делать, даже в таком возрасте, вроде согласились, но когда они встретили медсестру с нашего участка, она сказала, таааа вам не обязательно ее делать, куда вы там ходите и т. д и т. п. Хотя ходят и в магазин, и в поликлинику, и к ним приезжают гости, родные с других городов. Вопросов к вам у меня нет, но вот крик души из-за таких медсестер есть! Я всегда ЗА прививки, дочке всегда все делала по плану и она своей дочке так же все делает по плану.
Liudmyla
Вот и у меня появился вопрос. Можно ли как то достоверно определить уровень кальция в организме ребенка? Смотрела ваш выпуск где вы говорите о недостоверном показании кальция в крови. Дело в том, что у ребенка (1год и 8 месяцев) не закрыт родничек. На витамин Д3 высыпания в виде пятен на руках. Кальцикер не подходит тоже. Молочку не едим в любом виде. Я переживаю за то, что мало кальция у нас. Ребенок капризный, плохо спит. Зубы в хорошем состоянии 16 штук. Ногти ломаются, а на ногах ещё и ростут вверх. Успокойте маму и подскажите что делать или не делать!
Саша
Здравствуйте доктор! Меня интересует тема прикорма. В недавнем видео Вы сказали, что кое-какая информация по его введению изменилась. Сейчас нам 4 месяца. Начинать хотела с кефира в 6 месяцев. Но никто в моем окружении не начинает с кефира и врачи никому не говорят с него начинать. Мы здоровы, никаких проблем нет. Понимаю, не нужно смотреть, что делают другие. Но, все таки, хотелось бы узнать, актуально ли еще сегодня начинать прикорм с молочных продуктов по Вашей книге Здоровье ребенка. Заранее спасибо за ответ.
Мы здоровы, никаких проблем нет. Понимаю, не нужно смотреть, что делают другие. Но, все таки, хотелось бы узнать, актуально ли еще сегодня начинать прикорм с молочных продуктов по Вашей книге Здоровье ребенка. Заранее спасибо за ответ.
Надежда
Здравствуйте! Вопрос совсем не про болезни. Моей дочери 1, 7, у нее Очень хороший аппетит. Она есть много и все время голодна. Размер ее порций, примерно 200-250г, ест 4 раза в день каши, супы, творог, гарниры с мясом (сладости ест редко, чай пьет несладкий. Она с года ест сама ложкой, без мультиков. Рост у нее 87, вес 13, 5 кг. Она активная, я стараюсь с ней гулять 2 раза в день, чтобы больше ходила и энергию тратила. Врачи говорят, что все это из-за интенсивного роста, но я все равно волнуюсь. Страшно, что растолстеет.
Ksna
МНЕ ОЧЕНЬ НУЖЕН ВАШ СОВЕТ ( знаю это не по теме, но всё равно спрошу ). Смотрите, у меня с рождения есть кавернозная гемонгиома, на голиностопе. Ещё давно я смотрела вас по телевизору, вы там говорили, что к 14- ти годам она пройдёт, мне 13, а ничего не меняется.
БУДУ РАДА ЕСЛИ ВЫ ОТВЕТИТЕ. Меня давно мучает этот вопрос
Samsung
Евгений Олегович! На днях ходили на новую прививку. Медсестра вытащила из холодильника вакцину и, не встряхнув суспензию, собралась делать инъекцию ребенку! Я попросила ее хорошенько взболтать вакцину (прочитала об этом, изучая информацию в интернете. Влияет ли это на эффективность вакцины? Раньше на данный нюанс не обращала внимания, увы. На что еще обращать внимание в прививоч. кабинете? Спасибо
доктор Комаровский предупредил об опасной болезни
Читати українською
Медосмотр
Уля ЯдчукОб этом Евгений Олегович написал в своем Instagram. Читай, чтобы узнать подробнее.
В этот раз Евгений Олегович рассказал о частых причинах отита:
«В ухе образуется сера, и эта сера поддерживает в ухе совершенно определенную среду.
Если сера регулярно вымывается, например, водой (часто купаемся попадает вода или у тех, кто посещает бассейны), если в ухе регулярно ковыряются, если в ухе остаются комочки ваты, то все это провоцирует возникновение, так называемых, наружных отитов. Есть два микроба, которые вызывают наружный отит, оба со страшными названиями: стафилококк и синегнойная палочка.
Как правило, наружные отиты лечатся местно. Внутрь не глотается. Но лечатся, ничего тут страшного нет. Главное поймите: чем чаще вы ухаживаете за детскими ушами, тем больше риск развития наружного отита. То есть, как правило, если у ребенка в ухе что-то вскочило, то это не потому, что мама его не любит, а потому что мама любит его очень сильно». — заключил доктор.
Хуже бытовой химии: доктор Комаровский назвал самый опасный предмет в доме
Отит
Доктор Комаровский назвал лучший способ продления жизни пожилых родителейСмотри видео по теме:Наверное, все мы когда-то слышали историю о том, что чей-то дедушка всю жизнь пил крепленый портвейн и курил «Беломор», работал на вредном производстве, но при этом был абсолютно здоровым, а умер в возрасте 97 лет от несчастного случая. И в то же время слышим о крепком атлете тридцати лет, который регулярно тренируется, не вылезая из спортзала, придерживается здорового образа жизни и т.д., неожиданно заболевает раком. Почему так произошло? Ответ на этот вопрос дал авторитетный израильский онколог Цви Бернштейн на Первом форуме онкопациентов, который прошел 3 февраля в Киеве. Подробнее читай здесь.
И в то же время слышим о крепком атлете тридцати лет, который регулярно тренируется, не вылезая из спортзала, придерживается здорового образа жизни и т.д., неожиданно заболевает раком. Почему так произошло? Ответ на этот вопрос дал авторитетный израильский онколог Цви Бернштейн на Первом форуме онкопациентов, который прошел 3 февраля в Киеве. Подробнее читай здесь.
Напомним, доктор Комаровский — известный педиатр, «золотыми» правилами иммунитета которого пользуются все родители, неоднократно резко высказывал свое мнение о поведении слишком заботливых мамочек. Так, он заявил, что у родителей, которые «носятся» со своими детьми и содержат дом в идеальной чистоте, ребенок заболеет быстрее, чем у тех, кто относится к своим детям более равнодушно. Это высказывание вызвало бурю эмоций и обсуждений в обществе, но у Евгения Комаровского свое мнение на этот счет. Подробнее — читай, перейдя по ссылке.
Больше интересных материалов можно прочитать на clutch.ua.
Стиль Таюне: что любит носить «тайная жена» Олега Винника
Темы: доктор комаровский отит причины
Медосмотр
Как предотвратить пищевое отравление во время войны: 4 главных правила
Медосмотр
Первая медицинская помощь в Украине: что делать с сильными порезами кожи
Медосмотр
Климакс: гинеколог назвала ранние и поздние симптомы
Медосмотр
Высокий уровень холестерина: медики назвали симптомы, которые проявляются при ходьбе
Медосмотр
Как не заболеть раком: ТОП-7 рекомендаций врачей
Медосмотр
Гинеколог рассказала, какие анализы сдавать бесполезно: «Только при показаниях»
Медосмотр
Неалкогольная жировая болезнь печени: врачи назвали симптомы, которые указывают, что с печенью беда
Медосмотр
Боль в спине, усталость и не только: врачи назвали пять ранних симптомов рака почек
Медосмотр
Риск в 20-40 лет: гинеколог рассказала, что такое паратубарная киста
Медосмотр
Осторожно, глаукома: врачи назвали симптомы, которые указывают на развитие болезни
Читать дальше
Мелкоколонийные варианты Pseudomonas aeruginosa морщинистого типа избегают клиренса хозяина, являются гипервоспалительными и персистируют в различных средах хозяина
. 2 февраля 2018 г.; 14(2):e1006842.
2 февраля 2018 г.; 14(2):e1006842.
doi: 10.1371/journal.ppat.1006842. Электронная коллекция 2018, февраль.
Мэтью Дж. Пестрак 1 , Сара Б. Чейни 1 , Хизер С Эгглстон 1 , Шери Деллос-Нолан 1 , Сритея Диксит 2 , Шомита С Мэтью-Штайнер 2 , Сашвати Рой 2 , Мэтью Р. Парсек 3 , Чандан К Сен 2 , Даниэль Дж. Возняк 1
Принадлежности
- 1 Кафедра микробных инфекций и иммунитета, Университет штата Огайо, Колумбус, Огайо, Соединенные Штаты Америки.
- 2 Кафедра хирургии, Университет штата Огайо, Колумбус, Огайо, Соединенные Штаты Америки.
- 3 Кафедра микробиологии Вашингтонского университета, Сиэтл, Вашингтон, Соединенные Штаты Америки.
- PMID: 29394295
- PMCID: PMC5812653
- DOI: 10.1371/журнал.ppat.1006842
Бесплатная статья ЧВК
Мэтью Дж. Пестрак и др.
PLoS Патог. .
Бесплатная статья ЧВК
. 2 февраля 2018 г.; 14(2):e1006842.
doi: 10.1371/journal.ppat.1006842. Электронная коллекция 2018, февраль.
Авторы
Мэтью Дж. Пестрак 1 , Сара Б. Чейни 1 , Хизер С Эгглстон 1 , Шери Деллос-Нолан 1 , Сритея Диксит 2 , Шомита С Мэтью-Штайнер 2 , Сашвати Рой 2 , Мэтью Р. Парсек 3 , Чандан К Сен 2 , Даниэль Дж. Возняк 1
Возняк 1
Принадлежности
- 1 Кафедра микробных инфекций и иммунитета, Университет штата Огайо, Колумбус, Огайо, Соединенные Штаты Америки.
- 2 Кафедра хирургии, Университет штата Огайо, Колумбус, Огайо, Соединенные Штаты Америки.
- 3 Кафедра микробиологии Вашингтонского университета, Сиэтл, Вашингтон, Соединенные Штаты Америки.
- PMID: 29394295
- PMCID: PMC5812653
- DOI:
10.1371/журнал.
 ppat.1006842
ppat.1006842
Абстрактный
Pseudomonas aeruginosa вызывает разрушительные инфекции у людей с ослабленным иммунитетом. Однажды установленные инфекции P. aeruginosa становятся невероятно трудными для лечения из-за развития устойчивых к антибиотикам агрегированных сообществ, известных как биопленки. Клинический вариант P. aeruginosa, образующий гипербиопленку, известный как морщинистый вариант с небольшими колониями (RSCV), часто выделяют при хронических инфекциях и коррелируют с плохим клиническим исходом. Развитие этих мутантов во время инфекции предполагает избирательное преимущество этого фенотипа, но остается неясным, как этот фенотип способствует персистенции. В то время как предыдущие исследования предполагают, что RSCV могут выживать, уклоняясь от иммунного ответа хозяина, наше исследование показывает, что инфекция RSCV, PAO1ΔwspF, стимулирует обширный воспалительный ответ, который вызывает значительное повреждение окружающей ткани хозяина. Как в модели хронической раны, так и в модели острой легочной инфекции мы наблюдали повышенную бактериальную нагрузку, повреждение ткани хозяина и сильный ответ нейтрофилов во время инфекции RSCV. Учитывая важную роль нейтрофилов в заболевании, опосредованном P. aeruginosa, мы исследовали влияние фенотипа RSCV на функцию нейтрофилов. Фенотип RSCV способствует уклонению от фагоцитов и стимулирует выработку реактивных форм кислорода (АФК) нейтрофилами. Мы также демонстрируем, что агрегация бактерий и опосредованная TLR продукция провоспалительных цитокинов способствуют иммунному ответу на RSCV. Кроме того, RSCV проявляли повышенную толерантность к антимикробным препаратам, продуцируемым нейтрофилами, включая h3O2 и антимикробный пептид LL-37. В совокупности эти данные указывают на то, что RSCV вызывают сильный, но неэффективный ответ нейтрофилов, который вызывает значительное повреждение тканей хозяина. Это исследование дает новое представление о персистенции RSCV и указывает на то, что этот вариант может играть критическую роль в рецидивирующем повреждении тканей, часто связанном с хроническими инфекциями.
Как в модели хронической раны, так и в модели острой легочной инфекции мы наблюдали повышенную бактериальную нагрузку, повреждение ткани хозяина и сильный ответ нейтрофилов во время инфекции RSCV. Учитывая важную роль нейтрофилов в заболевании, опосредованном P. aeruginosa, мы исследовали влияние фенотипа RSCV на функцию нейтрофилов. Фенотип RSCV способствует уклонению от фагоцитов и стимулирует выработку реактивных форм кислорода (АФК) нейтрофилами. Мы также демонстрируем, что агрегация бактерий и опосредованная TLR продукция провоспалительных цитокинов способствуют иммунному ответу на RSCV. Кроме того, RSCV проявляли повышенную толерантность к антимикробным препаратам, продуцируемым нейтрофилами, включая h3O2 и антимикробный пептид LL-37. В совокупности эти данные указывают на то, что RSCV вызывают сильный, но неэффективный ответ нейтрофилов, который вызывает значительное повреждение тканей хозяина. Это исследование дает новое представление о персистенции RSCV и указывает на то, что этот вариант может играть критическую роль в рецидивирующем повреждении тканей, часто связанном с хроническими инфекциями.
Заявление о конфликте интересов
Авторы заявили об отсутствии конкурирующих интересов.
Цифры
Рис. 1. RSCV сохраняются во время моделирования…
Рис. 1. RSCV сохраняются во время модели хронической инфекции ожоговой раны и ингибируют рану…
Ожоги свиней были инфицированы через 3 дня PAO1 или PAO1Δ wspF . A) Биопсии ран были количественно определены для КОЕ. B и C) CLSM использовали для оценки бактериальной нагрузки. Р . aeruginosa окрашивали Alexa Fluor 488 (зеленый), а ткань хозяина — DAPI (синий). Количественно определяли площадь зеленой флуоресценции. Масштабные полосы указывают 35 мкм. D и E) Полоски ран были окрашены гематоксилин-эозином для оценки реэпителизации 35 d.i. Измеряли расстояние между эпителиальными язычками (ЭТ) в срезах тканей, окрашенных гематоксилин-эозином. Из-за ограничений размера H&E изображения на панели D были созданы путем слияния сканов двух разных слайдов, содержащих половину биопсии. Данные представлены как среднее ± SEM. *р<0,05, **р<0,01, ***р<0,001.
Р . aeruginosa окрашивали Alexa Fluor 488 (зеленый), а ткань хозяина — DAPI (синий). Количественно определяли площадь зеленой флуоресценции. Масштабные полосы указывают 35 мкм. D и E) Полоски ран были окрашены гематоксилин-эозином для оценки реэпителизации 35 d.i. Измеряли расстояние между эпителиальными язычками (ЭТ) в срезах тканей, окрашенных гематоксилин-эозином. Из-за ограничений размера H&E изображения на панели D были созданы путем слияния сканов двух разных слайдов, содержащих половину биопсии. Данные представлены как среднее ± SEM. *р<0,05, **р<0,01, ***р<0,001.
Рис. 2. RSCV уклоняются от фагоцитоза нейтрофилов.
А)…
Рис. 2. RSCV уклоняются от фагоцитоза нейтрофилов.
A) После заражения оценивали фагоцитоз нейтрофилов (MOI 1:50)…
Рис. 2. RSCV уклоняются от фагоцитоза нейтрофилов.
2. RSCV уклоняются от фагоцитоза нейтрофилов. A) Фагоцитоз нейтрофилов оценивали после инфицирования (MOI 1:50) с использованием CLSM. Нейтрофилы окрашивали DAPI (синий), внеклеточный Р . aeruginosa окрашивали Alexa Fluor 488 (зеленый) и нейтрофилы интернализовали P . aeruginosa окрашивали Alexa Fluor 647 (красный). Белые стрелки указывают на нейтрофилы, содержащие фагоцитированный P . синегнойная палочка . Б) Количественно определяли количество нейтрофилов, содержащих фагоцитированные P. aeruginosa , и определяли коэффициент интернализации. C) Фагоцитоз нейтрофилов оценивали с опсонизацией сыворотки или без нее путем количественного определения популяции, содержащей GFP, продуцирующий Р . aeruginosa после заражения (MOI 1:50). Данные представлены как среднее ± SEM. *р<0,05, **р<0,01.
Рис. 3. RSCV стимулируют выработку АФК нейтрофилами.
Рис. 3. RSCV стимулируют выработку АФК нейтрофилами.
А) Нейтрофилы были инфицированы логарифмическими бактериями…
Рис. 3. RSCV стимулируют выработку АФК нейтрофилами. A) Нейтрофилы инфицировали бактериями с логарифмической фазой (MOI 1:50) или обрабатывали PMA в присутствии репортера люминола. Люминесценцию измеряли в течение 60 минут (справа), площадь под кривой (AUC) рассчитывали и нормализовали по ответу PMA (слева). Изображения RLU репрезентативны для ответа ROS от нейтрофилов одного донора, измеренного в трех экземплярах, в то время как данные AUC были собраны с использованием нейтрофилов по крайней мере от 3 разных доноров. B и C) Ответ АФК на клинические изоляты RSCV МВ CF127 и CF39с. Данные представлены как среднее ± SEM. *п
*п
Рис. 4. Агрегация бактерий способствует АФК нейтрофилов…
Рис. 4. Агрегация бактерий способствует выработке АФК нейтрофилами, но одних экзополисахаридов недостаточно.
А)…
Рис. 4. Бактериальная агрегация способствует выработке АФК нейтрофилами, но одних экзополисахаридов недостаточно. A) ЭПС очищали от PAO1Δ pel P BAD -psl, PAO1Δ psl P BAD -pel или PAO1Δ pel Δ psl , а общее количество углеводов определяли с помощью серной кислоты. Уровни PAO1 или 10-кратные уровни PAO1 ePS добавляли к нейтрофилам человека, и ответ ROS измеряли с помощью репортера люминола. B) PAO1Δ psl Δ pel /pHERD20T и PAO1Δ psl Δ pel /pCdrAB выращивали до логарифмической фазы в присутствии 1% арабинозы, что приводило к образованию CdrA-опосредованных агрегатов в штамме, содержащем pCdrAB. Первичные нейтрофилы человека были инфицированы бактериями при МВД (1:50). C) Нейтрофилы обрабатывали супернатантом, собранным из культур PAO1/pCdrAB или PAO1∆9.0143 cdrA /pHERD20T и определяли количество АФК. Рассчитывали AUC ответа ROS в течение 1 часа и нормализовали по КОЕ инокуляционной культуры, чтобы обеспечить одинаковое количество клеток независимо от агрегации.
Первичные нейтрофилы человека были инфицированы бактериями при МВД (1:50). C) Нейтрофилы обрабатывали супернатантом, собранным из культур PAO1/pCdrAB или PAO1∆9.0143 cdrA /pHERD20T и определяли количество АФК. Рассчитывали AUC ответа ROS в течение 1 часа и нормализовали по КОЕ инокуляционной культуры, чтобы обеспечить одинаковое количество клеток независимо от агрегации.
Рис. 5. PAO1Δ wspF проявляет устойчивость к…
Рис. 5. PAO1Δ wspF проявляет толерантность к нейтрофильным противомикробным препаратам.
Лог-фазовые культуры PAO1 или…
Рис. 5. PAO1Δ wspF проявляет толерантность к антимикробным препаратам нейтрофилов. Культуры логарифмической фазы PAO1 или PAO1Δ wspF обрабатывали A) LL-37, B) H 2 O 2 и C) HOCl в меченых концентрациях в течение 15 мин. Количественно определяли КОЕ до и после лечения и определяли логарифмическую кратность уничтожения. Данные представлены как среднее ± SEM. ***р<0,001.
Количественно определяли КОЕ до и после лечения и определяли логарифмическую кратность уничтожения. Данные представлены как среднее ± SEM. ***р<0,001.
Рис. 6. D-PAO1Δ wspF гипервоспалительный и…
Рис. 6. D-PAO1Δ wspF является гипервоспалительным и сохраняется во время острой легочной инфекции.
Мыши были инфицированы…
Рис. 6. D-PAO1Δ wspF является гипервоспалительным и сохраняется при острой легочной инфекции. Мышей инфицировали бактериями 10 8 путем интраназальной инстилляции. Агрегаты PAO1Δ wspF разрушали иглой 22G перед инокуляцией, как указано (D-PAO1Δ wspF ). A) Легкие гомогенизировали и определяли количество КОЕ. B) Легкие окрашивали H&E 24 h. i. повреждение оценивалось как минимальное, среднее или серьезное. C) Во время инфекции PAO1Δ wspF крупные бактериальные агрегаты (черная пунктирная область) наблюдались через 2 часа в бронхиолах, а через 8 часов нейтрофильные пробки (желтая пунктирная область) наблюдались в бронхиолах. Шкала баров указывает 20 мкм. D) IL-1β и E) IL-6 в гомогенате легких измеряли с помощью ELISA. Данные представлены как среднее ± SEM. *р<0,05, **р<0,01.
i. повреждение оценивалось как минимальное, среднее или серьезное. C) Во время инфекции PAO1Δ wspF крупные бактериальные агрегаты (черная пунктирная область) наблюдались через 2 часа в бронхиолах, а через 8 часов нейтрофильные пробки (желтая пунктирная область) наблюдались в бронхиолах. Шкала баров указывает 20 мкм. D) IL-1β и E) IL-6 в гомогенате легких измеряли с помощью ELISA. Данные представлены как среднее ± SEM. *р<0,05, **р<0,01.
Рис. 7. D-PAO1Δ wspF легочная инфекция приводит…
Рис. 7. D-PAO1Δ wspF легочная инфекция приводит к тяжелому повреждению тканей и нейтрофильной инфильтрации по сравнению с…

Легкие мыши 24 л.с. окрашивали H&E для оценки нейтрофильной инфильтрации и повреждения легких на основе легочной архитектуры, некроза и гнойного воспаления. Изображения представляют легкие, обработанные или инфицированные A) PBS, B) PAO1, C) PAO1Δ wspF , D) D-PAO1Δ wspF . Масштабные полосы указывают 200 мкм.
Рис. 8. PAO1Δ wspF индуцирует провоспалительный цитокин…
Рис. 8. PAO1Δ wspF индуцирует продукцию провоспалительных цитокинов TLR-зависимым образом.
NR-9456 дикий тип и…
Рис. 8. PAO1Δ wspF индуцирует продукцию провоспалительных цитокинов TLR-зависимым образом. NR-9456 дикого типа и NR-15632 MyD88 -/- /TRIF -/- мышиные макрофаги были инфицированы PAO1 или PAO1Δ wspF на 4 часа. A) IL-1β и B) IL-6 измеряли в супернатантах клеток с помощью ELISA. ** р < 0,01. Данные представлены как среднее ± SEM.
A) IL-1β и B) IL-6 измеряли в супернатантах клеток с помощью ELISA. ** р < 0,01. Данные представлены как среднее ± SEM.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Межштаммовая динамика Pseudomonas aeruginosa и отбор мутантов гипербиопленки при хронической инфекции.
Глоаг Э.С., Маршалл К.В., Снайдер Д., Левин Г.Р., Харрис Дж.С., Сантос-Лопес А., Чейни С.Б., Уайтли М., Купер В.С., Возняк Д.Дж. Глоаг Э.С. и др. мБио. 201913 августа; 10(4):e01698-19. doi: 10.1128/mBio.01698-19. мБио. 2019. PMID: 31409682 Бесплатная статья ЧВК.
Варианты Pseudomonas aeruginosa морщинистых небольших колоний имеют адаптации, которые, вероятно, способствуют персистенции в легких при муковисцидозе.

Старки М., Хикман Дж. Х., Ма Л., Чжан Н., Де Лонг С., Хинц А., Паласиос С., Манойл С., Кириситс М.Дж., Старнер Т.Д., Возняк Д.Дж., Харвуд С.С., Парсек М.Р. Старки М. и др. J Бактериол. 2009 г.Июнь; 191 (11): 3492-503. doi: 10.1128/JB.00119-09. Epub 2009 27 марта. J Бактериол. 2009. PMID: 19329647 Бесплатная статья ЧВК.
Хроническая инфекция биопленки Pseudomonas aeruginosa повреждает мышиные S100A8/A9 и эффекторные цитокины нейтрофилов — последствия для отсроченного закрытия раны?
Трёструп Х., Лерче С.Дж., Кристоферсен Л.Дж., Томсен К., Дженсен П.О., Хоуген Х.П., Хойби Н., Мозер С. Trøstrup H, et al. Патог Дис. 2017 сен 29;75(7). doi: 10.1093/femspd/ftx068. Патог Дис. 2017. PMID: 28645160
Вариант с небольшими колониями складчатости и его гипербиопленка у Pseudomonas aeruginosa: адаптация, эволюция и биотехнологический потенциал.

Сюй А., Чжан С., Ван Т., Синь Ф., Ма Л.З., Чжоу Дж., Донг В., Цзян М. Сюй А и др. Биотехнология Adv. 2021 дек;53:107862. doi: 10.1016/j.biotechadv.2021.107862. Epub 2021 28 октября. Биотехнология Adv. 2021. PMID: 34718136 Обзор.
Иммунометаболиты дыхательных путей подпитывают инфекцию Pseudomonas aeruginosa.
Рикельме С.А., Принц А. Рикельме С.А. и др. Дыхание Рез. 2020 10 декабря; 21 (1): 326. doi: 10.1186/s12931-020-01591-x. Дыхание Рез. 2020. PMID: 33302964 Бесплатная статья ЧВК. Обзор.
Посмотреть все похожие статьи
Цитируется
Перспективы применения D-аминокислот при перипротезных инфекциях суставов.
Колдуэлл М., Хьюз М., Вэй Ф., Нго С., Паскуа Р., Пугаженди А.С., Коутап М.Дж. Колдуэлл М. и соавт. Кость рез. 2023 10 марта; 11 (1): 14. doi: 10.1038/s41413-023-00254-z. Кость рез. 2023. PMID: 36894568 Бесплатная статья ЧВК. Обзор.
Высокопроизводительное определение концентрации предохранителя от биопленки в течение Биопленки Pseudomonas aeruginosa с использованием синтетической среды для муковисцидоза мокроты.
Де Блеккер А., Ван ден Босше С., Де Саттер П.Дж., Бейренс Т., Краббе А., Коэнье Т. Де Блеккер А. и соавт. Биопленка. 2023 6 февраля; 5:100106. doi: 10.1016/j.bioflm.2023.100106. электронная коллекция 2023 дек. Биопленка. 2023. PMID: 36845825 Бесплатная статья ЧВК.
Влияние бикарбонатов на антибактериальный иммунитет и гликобиологию слизи при муковисцидозе легких: обзор с избранными экспериментальными наблюдениями.
Сью Р., Оу Т.Л., Дахеш С., Аконг К., Низет В. Сью Р. и соавт. Заразить микробы Дис. 2022 сен;4(3):103-110. doi: 10.1097/im9.0000000000000101. Epub 2022 16 августа. Заразить микробы Дис. 2022. PMID: 36793929 Бесплатная статья ЧВК.
Модуляция иммунного ответа клетками-персистерами Pseudomonas aeruginosa .
Гастингс CJ, Гиммлер GE, Патель А, Маркес CNH. Hastings CJ и др. bioRxiv. 2023 8 января: 2023.01.07.523056. дои: 10.1101/2023.01.07.523056. Препринт. bioRxiv. 2023. PMID: 36711557 Бесплатная статья ЧВК. Обновлено.
Разработка имплантируемой трехмерной модели функциональной патогенной многовидовой биопленки для изучения инфицированных ран.

Карденас-Кальдерон К., Велосо-Хименес В., Гонсалес Т., Возняк А., Гарсия П., Мартин С.С., Варас Х.Ф., Карраско-Вонг И., Вера М., Эганья Х.Т. Карденас-Кальдерон С. и др. Научный представитель 2022 г. 17 декабря; 12 (1): 21846. doi: 10.1038/s41598-022-25569-5. Научный представитель 2022. PMID: 36528648 Бесплатная статья ЧВК.
Просмотреть все статьи «Цитируется по»
Рекомендации
- Магилл С.С., Эдвардс Дж.Р., Бамберг В., Белдавс З.Г., Думяти Г., Кайнер М.а. и соавт. Многострановое точечное обследование распространенности инфекций, связанных с оказанием медицинской помощи. N Engl J Med. 2014; 1198–208. — ЧВК — пабмед
- Вайнштейн М.
 П., Таунс М.Л., Куарти С.М., Мирретт С., Реймер Л.Г., Пармиджани Г., Реллер Л. Клиническое значение положительных культур крови в 1990-х годах: проспективная комплексная оценка микробиологии, эпидемиологии и исходов бактериемии и фунгемии у взрослых . Клин Инфекция Дис. 1997; 584–602.
—
пабмед
П., Таунс М.Л., Куарти С.М., Мирретт С., Реймер Л.Г., Пармиджани Г., Реллер Л. Клиническое значение положительных культур крови в 1990-х годах: проспективная комплексная оценка микробиологии, эпидемиологии и исходов бактериемии и фунгемии у взрослых . Клин Инфекция Дис. 1997; 584–602.
—
пабмед
- Вайнштейн М.
- Костертон Дж. В., Стюарт П. С., Гринберг Э. П. Бактериальные биопленки: частая причина стойких инфекций. 1999; 1318–23. — пабмед
- Малкахи Л.Р., Изабелла В.М., Льюис К.
 Биопленки Pseudomonas aeruginosa при заболеваниях. Микроб Экол. 2013;1–12.
—
ЧВК
—
пабмед
Биопленки Pseudomonas aeruginosa при заболеваниях. Микроб Экол. 2013;1–12.
—
ЧВК
—
пабмед
- Малкахи Л.Р., Изабелла В.М., Льюис К.
- Høiby N. Последние достижения в лечении инфекций Pseudomonas aeruginosa при муковисцидозе. БМС Мед. 2011; 32 doi: 10.1186/1741-7015-9-32 — DOI — ЧВК — пабмед
Типы публикаций
термины MeSH
вещества
Грантовая поддержка
- Р33 АИ119116/AI/NIAID NIH HHS/США
- P30 CA016058 /NH/NIH HHS/США
- R01 AI077628/AI/NIAID NIH HHS/США
- NR013898 /NH/NIH HHS/США
- NR015676 /NH/NIH HHS/США
- T32 GM068412/GM/NIGMS NIH HHS/США
- AI119116 /NH/NIH HHS/США
- R01 NR015676/NR/NINR NIH HHS/США
- R01 AI097511/AI/NIAID NIH HHS/США
- R21 AI119116/AI/NIAID NIH HHS/США
- R01 NR013898/NR/NINR NIH HHS/США
- P30 CA016058/CA/NCI NIH HHS/США
- AI097511 /NH/NIH HHS/США
Полнотекстовые ссылки
Публичная научная библиотека Бесплатная статья ЧВК
Укажите
Формат: ААД АПА МДА НЛМ
Отправить на
Регуляция конъюгативного переноса плазмид и интегративных конъюгативных элементов
8 Цитаты в Европе PMC
Дата публикации в Европе PubMed Central.
Регуляция грамположительной конъюгации.
Колер В., Келлер В., Грохманн Э., Front Microbiol 10(), 2019
PMID: 31191478
Гены, стимулирующие рост растений, могут превращаться в факторы вирулентности посредством горизонтального переноса генов.
Штритцлер М., Сото Г., Аюб Н., Microb Ecol 76(3), 2018 г.
PMID: 29476343
Лекарственная устойчивость и механизмы переноса генов в респираторных/оральных бактериях.
Цзян С., Цзэн Дж., Чжоу С., Ли Ю., Дж. Дент Рез 97(10), 2018 г.
PMID: 29928825
Интегративный конъюгативный элемент clc (ICEclc) Pseudomonas aeruginosa JB2.
Оби К.С., Вайла С., де Ганнес В., Беррес М.Е., Уокер Дж., Павелек Д., Хайман Дж., Хикки В.Дж., Front Microbiol 9(), 2018 г.
PMID: 30050515
TraN: Новый репрессор конъюгированной системы секреции Enterococcus IV типа.
Kohler V, Goessweiner-Mohr N, Aufschnaiter A, Fercher C, Probst I, Pavkov-Keller T, Hunger K, Wolinski H, Büttner S, Grohmann E, Keller W. , Nucleic Acids Res 46(17), 2018
, Nucleic Acids Res 46(17), 2018
PMID: 30060171
Стратегии борьбы с устойчивостью к противомикробным препаратам: антиплазмидное и плазмидное лечение.
Buckner MMC, Ciusa ML, Piddock LJV., FEMS Microbiol Rev 42(6), 2018 г.
PMID: 30085063
Появление доминирующих групп бактерий с множественной лекарственной устойчивостью: уроки истории и полногеномного секвенирования.
Клемм Э.Дж., Вонг В.К., Дуган Г., Proc Natl Acad Sci U S A 115(51), 2018
PMID: 30559200
Эволюция стабильности плазмид: являются ли инфекционная передача и компенсаторная эволюция конкурирующими эволюционными траекториями?
Hall JPJ, Brockhurst MA, Dytham C, Harrison E., Плазмида 91(), 2017 г.
PMID: 28461121
110 Ссылки
Daten bereitgestellt von Europe PubMed Central.
Биологическое разнообразие прокариотических систем секреции IV типа.
Alvarez-Martinez CE, Christie PJ., Microbiol. Мол. биол. Ред. 73(4), 2009 г.
Мол. биол. Ред. 73(4), 2009 г.
PMID: 19946141
Горизонтальный обмен генами в микробиоте окружающей среды.
Аминов Р.И., Фронт Микробиол 2(), 2011
PMID: 21845185
Биология ICEBs1, интегративного и конъюгативного элемента Bacillus subtilis.
Auchtung JM, Алексанян Н, Булку А, Беркмен МБ., Plasmid 86(), 2016
PMID: 27381852
Горизонтальный перенос плазмиды устойчивости и вирулентности Salmonella enterica Serovar Infantis pESI в кишечную микробиоту теплокровных хозяев.
Авив Г., Рахав Г., Гал-Мор О., MBio 7(5), 2016 г.
PMID: 27601577
Функциональная идентификация областей конъюгации и репликации плазмиды устойчивости к тетрациклину pCW3 из Clostridium perfringens.
Bannam TL, Teng WL, Bulach D, Lyras D, Rood JI., J. Bacteriol. 188(13), 2006 г.
PMID: 16788202
Антибиотикорезистентность, формирующая многоуровневую популяционную биологию бактерий.
Baquero F, Tedim AP, Coque TM., Front Microbiol 4(), 2013 г.
PMID: 23508522
Последовательность генома энтеробактериального фитопатогена Erwinia carotovora subsp. atroseptica и характеристика факторов вирулентности.
Белл К.С., Себайхиа М., Причард Л., Холден М.Т., Хайман Л.Дж., Холева М.С., Томсон Н.Р., Бентли С.Д., Черчер Л.Дж., Мангалл К., Аткин Р., Бейсон Н., Брукс К., Чиллингворт Т., Кларк К., Доггетт Дж., Фрейзер A, Hance Z, Hauser H, Jagels K, Moule S, Norbertczak H, Ormond D, Price C, Quail MA, Sanders M, Walker D, Whitehead S, Salmond GP, Birch PR, Parkhill J, Toth IK., Проц. Натл. акад. науч. США 101(30), 2004 г.
PMID: 15263089
pSymA-зависимая мобилизация мегаплазмиды Sinorhizobium meliloti pSymB.
Бланка-Ордонез Х., Олива-Гарсия Дж. Дж., Перес-Мендоса Д., Сото М. Дж., Оливарес Дж., Санхуан Дж., Ногалес Дж., Дж. Бактериол. 192(23), 2010 г.
PMID: 20889746
Регуляция горизонтального переноса генов в Bacillus subtilis путем активации консервативной сайт-специфичной протеазы.
Бозе Б, Гроссман А. Д., J. Бактериол. 193(1), 2010 г.
PMID: 21036995
У Rhizobium etli симбиотический перенос плазмиды, конкурентность клубеньков и клеточный рост требуют взаимодействия между различными репликонами.
Бром С., Гарсия-де лос Сантос А., Сервантес Л., Паласиос Р., Ромеро Д., Плазмида 44(1), 2000 г.
PMID: 10873525
Перенос симбиотической плазмиды Rhizobium etli CFN42 требует коинтеграции с p42a, которая может быть опосредована сайт-специфической рекомбинацией.
Бром С., Жирар Л., Тун-Гарридо С., Гарсия-де-лос-Сантос А., Бустос П., Гонсалес В., Ромеро Д., J. Bacteriol. 186(22), 2004 г.
PMID: 15516565
Границы конъюгативного переноса ризобиальных плазмид: сдерживающие и высвобождающие факторы
Brom, 2014
Конъюгальный перенос плазмиды вирулентности Salmonella enterica регулируется лейцин-чувствительным регуляторным белком и адениновым метилированием ДНК.
Camacho EM, Casadesus J. , Mol. микробиол. 44(6), 2002 г.
, Mol. микробиол. 44(6), 2002 г.
PMID: 12067346
Регуляция конъюгального переноса путем метилирования Lrp и Dam в плазмиде R100.
Камачо Э.М., Серна А., Касадесус Дж., Междунар. микробиол. 8(4), 2005 г.
PMID: 16562380
Распутывание регуляторной сети мобилизации плазмиды IncA/C: когда геномные островки захватывают конъюгативные элементы.
Карраро Н., Матто Д., Буррус В., Родриг С., Mob Genet Elements 5(3), 2015 г.
PMID: 26442183
Конъюгативные плазмиды IncA/C мобилизуют новое семейство островков множественной лекарственной устойчивости в клинических изолятах Vibrio cholerae Non-O1/Non-O139 из Гаити.
Карраро Н., Ривард Н., Чеккарелли Д., Колвелл Р.Р., Буррус В., MBio 7(4), 2016 г.
PMID: 27435459
Островок патогенности Pseudomonas aeruginosa PAPI-1 передается через пилюс нового типа IV.
Carter MQ, Chen J, Lory S., J. Bacteriol. 192(13), 2010 г.
PMID: 20363934
Распространение Tn916 требуется для экспрессии функций передачи, кодируемых транспозонами: характеристика длинных индуцируемых тетрациклином транскриптов, считываемых через сайт прикрепления.
Celli J, Trieu-Cuot P., Mol. микробиол. 28(1), 1998 г.
PMID: 9593300
Использование эксцизионной репортерной плазмиды для изучения внутриклеточной подвижности конъюгативного транспозона Tn916 у грамположительных бактерий.
Celli J, Poyart C, Trieu-Cuot P., Microbiology (Reading, Engl.) 143 (Pt 4)(), 1997
PMID: 9141688
Конъюгативная плазмида клубенькового штамма Sinorhizobium fredii, образующего фасоль, собрана из последовательностей двух плазмид Rhizobium и хромосомы штамма Sinorhizobium.
Сервантес Л., Бустос П., Жирар Л., Сантамария Р.И., Давила Г., Винуэса П., Ромеро Д., Бром С., BMC Microbiol. 11(), 2011 г.
PMID: 21702991
Идентификация генов, необходимых для вырезания CTnDOT, конъюгативного транспозона Bacteroides.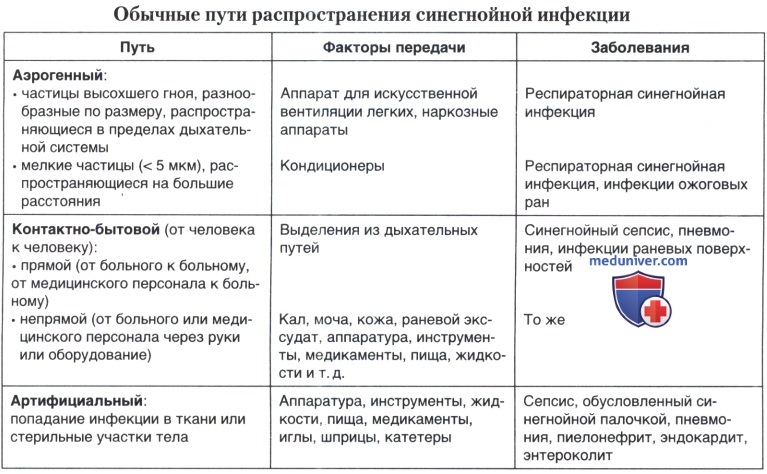
Ченг К., Сутанто Ю., Шумейкер Н.Б., Гарднер Дж.Ф., Сальерс А.А., Мол. микробиол. 41(3), 2001 г.
PMID: 11532130
Транссексуальность в ризосфере: определение кворума обратимо превращает Agrobacterium tumefaciens из фенотипически женского в мужской.
Cho H, Pinto UM, Winans SC., J. Bacteriol. 191(10), 2009 г.
PMID: 19304847
Передача сигналов феромона пептида в Streptococcus и Enterococcus.
Cook LC, Federle MJ., FEMS Microbiol. Ред. 38(3), 2013 г.
PMID: 24118108
Индуцированный реципиентом перенос симбиотической плазмиды pRL1JI в Rhizobium leguminosarum bv. viciae регулируется датчиком кворума.
Danino VE, Wilkinson A, Edwards A, Downie JA., Mol. микробиол. 50(2), 2003 г.
PMID: 14617175
Влияние распространения плазмид деградации 2,4-дихлорфеноксиуксусной кислоты (2,4-D) на деградацию 2,4-D и на структуру бактериального сообщества в двух разных почвенных горизонтах.
Деджонге В., Горис Дж., Эль Фантрусси С., Хофте М., Де Вос П., Верстрате В., Топ EM., Заявл. Окружающая среда. микробиол. 66(8), 2000 г.
PMID: 10919784
Системы переноса плазмид в ризобиях.
Дин Х., Хайнс М.Ф., Кан. Дж. Микробиол. 55(8), 2009 г.
PMID: 19898531
Генетическая характеристика новой системы конъюгации ризобиальных плазмид у Rhizobium leguminosarum bv. viciae штамм VF39SM.
Ding H, Yip CB, Hynes MF., J. Bacteriol. 195(2), 2012 г.
PMID: 23144250
Свойства шести плазмид деградации пестицидов, выделенных из Alcaligenes paradoxus и Alcaligenes eutrophus.
Don RH, Pemberton JM., J. Bacteriol. 145(2), 1981 г.
PMID: 6257648
Пептид-феромон-индуцируемая система конъюгации плазмиды pCF10 Enterococcus faecalis: межклеточная передача сигналов, перенос генов, сложность и эволюция.
Данни GM., Филос. Транс. Р. Соц. Lond., B, Biol. науч. 362(1483), 2007 г.
PMID: 17360276
Энтерококковые половые феромоны: эволюционные пути к сложным двухсигнальным системам.
Данни ГМ, Бернтссон Р.П., J. Бактериол. 198(11), 2016
PMID: 27021562
Индуцированная агрегация клеток и спаривание у Streptococcus faecalis: свидетельство существования бактериального полового феромона.
Dunny GM, Brown BL, Clewell DB., Proc. Натл. акад. науч. США 75(7), 1978 г.
PMID: 98769
Ингибирование и активация синтеза бактериальной люциферазы.
Эберхард А., Дж. Бактериол. 109(3), 1972 г.
PMID: 5011244
Совместное распространение устойчивости к металлам и антибиотикам в плазмидах ST3-IncHI2 из изолятов E. coli от сельскохозяйственных животных.
Fang L, Li X, Li L, Li S, Liao X, Sun J, Liu Y., Sci Rep 6(), 2016
PMID: 27143648
Мобильные генетические элементы: агенты эволюции с открытым исходным кодом.
Frost LS, Leplae R, Summers AO, Toussaint A. , Nat. Преподобный Микробиолог. 3(9), 2005 г.
, Nat. Преподобный Микробиолог. 3(9), 2005 г.
PMID: 16138100
ГЕНЫ АГРОБАКТЕРИЙ И РАСТЕНИЙ, УЧАСТВУЮЩИХ В ПЕРЕНОСЕ И ИНТЕГРАЦИИ Т-ДНК.
Гельвин С.Б., Анну. Преподобный Завод Физиол. Завод Мол. биол. 51(), 2000 г.
PMID: 15012192
Конъюгация в грамположительных бактериях.
Гессвайнер-Мор Н., Арендс К., Келлер В., Грохманн Э., Microbiol Spectr 2(4), 2014 г.
PMID: 26104193
Реакция растений на Agrobacterium tumefaciens и развитие корончатого галла.
Гольке Дж., Дикен Р., Front Plant Sci 5(), 2014 г.
PMID: 24795740
Консервативные последовательности симбиотической плазмидной ДНК в мультирепликонной пангеномной структуре Rhizobium etli.
Гонсалес В., Акоста Х.Л., Сантамария Р.И., Бустос П., Фернандес Х.Л., Эрнандес Гонсалес И.Л., Диас Р., Флорес М., Паласиос Р., Мора Х., Давила Г., Заявл. Окружающая среда. микробиол. 76(5), 2010
PMID: 20048063
Положительный регулятор TraJ плазмиды F Escherichia coli нестабилен на фоне cpxA*.
Губбинс М.Дж., Лау И., Уилл В.Р., Манчак Дж.М., Райвио Т.Л., Фрост Л.С., Дж. Бактериол. 184(20), 2002 г.
PMID: 12270837
Функциональный анализ детерминанты устойчивости к бацитрацину, расположенной на ICECp1, новом Tn916-подобном элементе из конъюгативной плазмиды Clostridium perfringens.
Хан X, Ду XD, Саути Л., Булах Д.М., Зееманн Т., Ян XX, Баннам Т.Л., Руд Дж.И., Антимикроб. Агенты Чемотер. 59(11), 2015
PMID: 26282424
Сборка и перенос трехкомпонентных интегративных и конъюгативных генетических элементов.
Haskett TL, Terpolilli JJ, Bekuma A, O’Hara GW, Sullivan JT, Wang P, Ronson CW, Ramsay JP., Proc. Натл. акад. науч. США 113(43), 2016 г.
PMID: 27733511
Чувство кворума у Rhizobium sp. штамм NGR234 регулирует экспрессию генов конъюгального переноса (tra) и влияет на скорость роста.
Хе Х, Чанг В., Пирс Д.Л., Сейб Л.О., Вагнер Дж., Фукуа К., J. Бактериол. 185(3), 2003 г.
PMID: 12533456
Плазмиды способствуют диверсификации и адаптации популяций бактерий в почве.
Heuer H, Smalla K., FEMS Microbiol. Ред. 36(6), 2012 г.
PMID: 22393901
Сеть передачи бактериальных мобильных резистомов, соединяющая микробиомы животных и человека.
Hu Y, Yang X, Li J, Lv N, Liu F, Wu J, Lin IY, Wu N, Weimer BC, Gao GF, Liu Y, Zhu B., Appl. Окружающая среда. микробиол. 82(22), 2016
PMID: 27613679
Интегративные и сопряженные элементы (ICE): что они делают и как они работают.
Johnson CM, Grossman AD., Annu. Преподобный Жене. 49(), 2015 г.
PMID: 26473380
Как ризобиальные симбионты вторгаются в растения: модель Sinorhizobium-Medicago.
Джонс К.М., Кобаяши Х., Дэвис Б.В., Тага М.Е., Уокер Г.К., Нац. Преподобный Микробиолог. 5(8), 2007 г.
PMID: 17632573
Исторический отчет о понимании механизма онкогенеза корончатых галлов, индуцированного Agrobacterium tumefaciens.
Kado CI., Front Microbiol 5(), 2014 г.
PMID: 25147542
Плазмиды pJP4 и r68.45 могут переноситься между популяциями Bradyrhizobia в нестерильной почве.
Kinkle BK, Sadowsky MJ, Schmidt EL, Koskinen WC., Appl. Окружающая среда. микробиол. 59(6), 1993 г.
PMID: 16348953
Металлические стрессоры последовательно модулируют потенциал поглощения бактериальной конъюгальной плазмиды филогенетически консервативным образом.
Клумпер Ю., Дечесне А., Рибер Л., Брандт К.К., Гулай А., Соренсен С.Дж., Сметс Б.Ф., ISME J 11(1), 2016 г.
PMID: 27482924
Горизонтальный перенос генов: сущность и эволюционность у прокариот и роль в эволюционных переходах
Koonin, 2016
Социальное поведение и принятие решений при бактериальной конъюгации.
Koraimann G, Wagner MA., Front Cell Infect Microbiol 4(), 2014 г.
PMID: 24809026
Глобальная транскрипционная регуляция генов остова в плазмиде RA3 с широким диапазоном хозяев из группы IncU включает сегрегационный белок KorB (семейство ParB).
Кулинска А., Годзишевская Ю., Войцеховская А., Людвичак М., Ягура-Бурджи Г., Заяв. Окружающая среда. микробиол. 82(8), 2016
PMID: 26850301
Плазмиды с хромосомоподобной ролью у ризобий.
Landeta C, Davalos A, Cevallos MA, Geiger O, Brom S, Romero D., J. Bacteriol. 193(6), 2011 г.
PMID: 21217003
Функции и регуляция чувства кворума у Agrobacterium tumefaciens.
Ланг Дж., Фор Д., Front Plant Sci 5(), 2014 г.
PMID: 24550924
Токсиновые плазмиды Clostridium perfringens.
Li J, Adams V, Bannam TL, Miyamoto K, Garcia JP, Uzal FA, Rood JI, McClane BA., Microbiol. Мол. биол. Ред. 77(2), 2013 г.
PMID: 23699255
Индукторы образования клубеньков растений усиливают горизонтальный перенос генов островного симбиоза Azorhizobium caulinodans.
Линг Дж., Ван Х., Ву П., Ли Т., Тан Ю., Насир Н., Чжэн Х., Массон-Бойвин С., Чжун З., Чжу Дж., Proc. Натл. акад.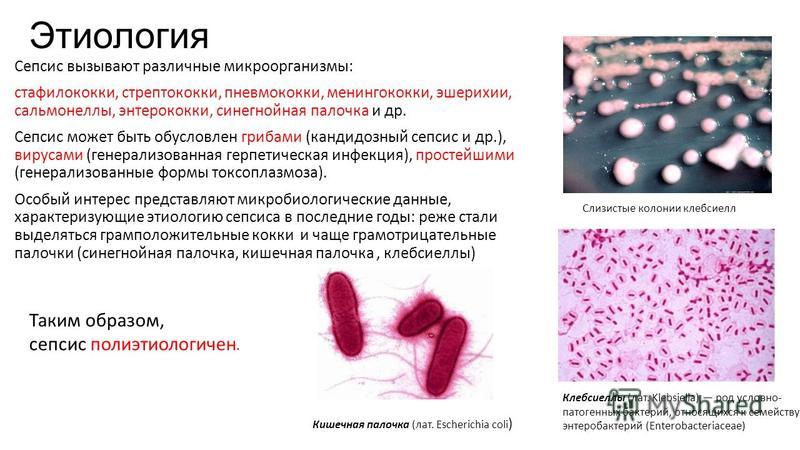 науч. США 113(48), 2016 г.
науч. США 113(48), 2016 г.
PMID: 27849579
Эволюционные пути, расширяющие диапазон плазмидных хозяев: последствия для распространения устойчивости к антибиотикам.
Loftie-Eaton W, Yano H, Burleigh S, Simmons RS, Hughes JM, Rogers LM, Hunter SS, Settles ML, Forney LJ, Ponciano JM, Top EM., Mol. биол. Эвол. 33(4), 2015
PMID: 26668183
Гены, кодирующие консервативные гипотетические белки, локализованные в области конъюгативного переноса плазмиды pRet42a из Rhizobium etli CFN42, участвуют в модуляции переноса и влияют на конъюгацию от разных доноров
Лопес-Фуэнтес, Фронт. микробиол. 5(), 2015
Глобальный регулятор транскрипции KorC координирует экспрессию трех основных модулей плазмиды RA3 широкого круга хозяев из группы несовместимости IncU.
Ludwiczak M, Dolowy P, Markowska A, Szarlak J, Kulinska A, Jagura-Burdzy G., Plasmid 70(1), 2013
PMID: 23583562
Неинвазивное определение конъюгативного переноса плазмид, несущих гены устойчивости к антибиотикам, в бактериях, связанных с биопленкой: влияние загрузки субстрата и выбора антибиотика.
Ma H, Bryers JD., Appl. микробиол. Биотехнолог. 97(1), 2012 г.
PMID: 22669634
Регулируемая определением кворума инициация транскрипции генов переноса и репликации плазмид в биоваре Rhizobium leguminosarum viciae.
McAnulla C, Edwards A, Sanchez-Contreras M, Sawers RG, Downie JA., Microbiology (Reading, Engl.) 153 (Pt 7), 2007 г.
PMID: 17600052
Анализ транскриптома показал, что система распознавания кворума регулирует перенос мегаплазмиды pAt в Agrobacterium tumefaciens.
Мхедби-Хаджри Н., Яхиауи Н., Монди С., Хью Н., Пелисье Ф., Фор Д., Дессо Ю., BMC Genomics 17(), 2016
PMID: 27543103
Плазмиды pMOL28 и pMOL30 Cupriavidus metallidurans специализируются на максимально жизнеспособном ответе на тяжелые металлы.
Monchy S, Benotmane MA, Janssen P, Vallaeys T, Taghavi S, van der Lelie D, Mergeay M., J. Bacteriol. 189(20), 2007 г.
PMID: 17675385
Клеточный контроль синтеза и активности люминесцентной системы бактерий.
Nealson KH, Platt T, Hastings JW., J. Bacteriol. 104(1), 1970 г.
PMID: 5473898
Конъюгальный перенос симбиотической плазмиды Sinorhizobium meliloti 1021 регулируется согласованным действием одно- и двухкомпонентных регуляторов передачи сигнала.
Ногалес Дж., Бланка-Ордоньес Х., Оливарес Дж., Санхуан Дж., Окружающая среда. микробиол. 15(3), 2013 г.
PMID: 23336126
Последовательности источника переноса облегчают мобилизацию неконъюгативных плазмид устойчивости к противомикробным препаратам в Staphylococcus aureus.
O’Brien FG, Yui Eto K, Murphy RJ, Fairhurst HM, Coombs GW, Grubb WB, Ramsay JP., Nucleic Acids Res. 43(16), 2015
PMID: 26243776
Боковой перенос генов и природа бактериальных инноваций.
Очман Х., Лоуренс Дж.Г., Гройсман Э.А., Природа 405(6784), 2000
PMID: 10830951
Конъюгальный перенос генов деградации полихлорированного бифенила/бифенила в Acidovorax sp. штамм ККС102, которые расположены по интегративному и конъюгативному элементам.
штамм ККС102, которые расположены по интегративному и конъюгативному элементам.
Оцубо Ю., Исибаши Ю., Наганава Х., Хирокава С., Атобе С., Нагата Ю., Цуда М., J. Bacteriol. 194(16), 2012 г.
PMID: 22685277
Идентификация гена rctA, необходимого для репрессии конъюгативного переноса ризобиальных симбиотических мегаплазмид.
Перес-Мендоса Д., Сепульведа Э., Пандо В., Муньос С., Ногалес Дж., Оливарес Дж., Сото М.Дж., Эррера-Сервера Дж.А., Ромеро Д., Бром С., Санхуан Дж., Дж. Бактериол. 187(21), 2005 г.
PMID: 16237017
Фактор конъюгации Agrobacterium tumefaciens регулирует перенос плазмиды Ti путем аутоиндукции.
Piper KR, Beck von Bodman S, Farrand SK., Nature 362(6419), 1993
PMID: 8464476
Идентификация трансмиссивной плазмиды из аргентинского штамма Sinorhizobium meliloti, которая может быть мобилизована конъюгативными хелперными функциями европейского штамма S. meliloti GR4.
Писторио М. , Дель Папа М.Ф., Балаге Л.Дж., Лагарес А., FEMS Microbiol. лат. 225(1), 2003 г.
, Дель Папа М.Ф., Балаге Л.Дж., Лагарес А., FEMS Microbiol. лат. 225(1), 2003 г.
PMID: 125
rptA, новый ген из Ensifer (Sinorhizobium) meliloti, участвующий в супружеском переносе.
Писторио М., Торрес Техеризо Г.А., Дель Папа М.Ф., Джусти Мде Л., Лосано М., Лагарес А., FEMS Microbiol. лат. 345(1), 2013 г.
PMID: 23672494
Расширенные регуляторные сети интегративных и конъюгативных элементов SXT/R391 и конъюгативных плазмид IncA/C.
Poulin-Laprade D, Carraro N, Burrus V., Front Microbiol 6(), 2015 г.
PMID: 26347724
Межштаммовой перенос большого острова патогенности (PAPI-1) Pseudomonas aeruginosa.
Цю X, Гуркар AU, Лори С., Proc. Натл. акад. науч. США 103(52), 2006 г.
PMID: 17179047
Вырезание и перенос симбиозного островка Mesorhizobium loti R7A требует интегразы IntS, нового фактора направленности рекомбинации RdfS и предполагаемой релаксазы RlxS.
Ramsay JP, Sullivan JT, Stuart GS, Lamont IL, Ronson CW., Mol. микробиол. 62(3), 2006 г.
PMID: 17076666
Регуляторная система семейства LuxRI контролирует вырезание и перенос симбиотического островка штамма Mesorhizobium loti R7A путем активации экспрессии двух консервативных гипотетических генов.
Ramsay JP, Sullivan JT, Jambari N, Ortori CA, Heeb S, Williams P, Barrett DA, Lamont IL, Ronson CW., Mol. микробиол. 73(6), 2009 г.
PMID: 19682258
Широко распространенный молекулярный переключатель контролирует кворумное восприятие и перенос островков симбиоза у Mesorhizobium loti посредством экспрессии нового антиактиватора.
Ramsay JP, Major AS, Komarovsky VM, Sullivan JT, Dy RL, Hynes MF, Salmond GP, Ronson CW., Mol. микробиол. 87(1), 2012 г.
PMID: 23106190
Обновленный взгляд на конъюгацию и мобилизацию плазмид у Staphylococcus.
Ramsay JP, Kwong SM, Murphy RJ, Yui Eto K, Price KJ, Nguyen QT, O’Brien FG, Grubb WB, Coombs GW, Firth N. , Mob Genet Elements 6(4), 2016
, Mob Genet Elements 6(4), 2016
PMID: 27583185
Низкочастотный горизонтальный перенос элемента, содержащего гены деградации хлоркатехола из Pseudomonas sp. штамм B13 к Pseudomonas putida F1 и местным бактериям в лабораторных микрокосмах с активным илом.
Ravatn R, Zehnder AJ, van der Meer JR., Appl. Окружающая среда. микробиол. 64(6), 1998 г.
PMID: 9603824
Romero, 2004
Необычная экспрессия гена интегразы на геномном островке clc у Pseudomonas sp. штамм В13.
Sentchilo V, Ravatn R, Werlen C, Zehnder AJ, van der Meer JR., J. Bacteriol. 185(15), 2003 г.
PMID: 12867462
Характеристика двух альтернативных промоторов для экспрессии интегразы в геномном островке clc Pseudomonas sp. штамм В13.
Sentchilo V, Zehnder AJ, van der Meer JR., Mol. микробиол. 49(1), 2003 г.
PMID: 12823813
Транскрипционная интерференция и репрессия модулируют конъюгативную способность симбиотической плазмиды Rhizobium etli.
Сепульведа Э., Перес-Мендоса Д., Рамирес-Ромеро М.А., Сото М.Дж., Лопес-Лара И.М., Гейгер О., Санхуан Дж., Бром С., Ромеро Д., J. Bacteriol. 190(12), 2008 г.
PMID: 18424522
Белок-сегрегатор хромосом превратился в конъюгативный белок-транслокатор ДНК.
Сепульведа Э., Фогельманн Дж., Мут Г., Mob Genet Elements 1(3), 2011 г.
PMID: 22479692
Тетрациклин усиливает опосредованный Tn916 конъюгальный перенос.
Showsh SA, Andrews RE Jr., Plasmid 28(3), 1992 г.
PMID: 1334267
Введение плазмиды в подповерхностные микрокосмы, подверженные стрессу металлов: судьба плазмиды и реакция сообщества.
Сметс Б.Ф., Морроу Дж.Б., Аранго Пинедо К., Заяв. Окружающая среда. микробиол. 69(7), 2003 г.
PMID: 12839785
Реакция Agrobacterium tumefaciens на сигнальные молекулы растительного происхождения.
Субрамони С., Натху Н., Климов Э., Юань З.С., Front Plant Sci 5(), 2014 г.
PMID: 25071805
Эволюция ризобий путем приобретения симбиозного острова размером 500 т.п.н., который интегрируется в ген phe-тРНК.
Салливан Дж.Т., Ронсон К.В., Proc. Натл. акад. науч. США 95(9), 1998 г.
PMID: 9560243
Клубеньковые штаммы Rhizobium loti возникают в результате хромосомного симбиотического переноса генов в окружающей среде.
Салливан Дж.Т., Патрик Х.Н., Лоутер В.Л., Скотт Д.Б., Ронсон К.В., Proc. Натл. акад. науч. США 92(19), 1995 г.
PMID: 7568057
Ртуть, выделяющаяся из зубных «серебряных» пломб, провоцирует рост устойчивых к ртути и антибиотикам бактерий в ротовой и кишечной флоре приматов.
Саммерс А.О., Уайрман Дж., Вими М.Дж., Лоршайдер Ф.Л., Маршалл Б., Леви С.Б., Беннетт С., Биллард Л., Антимикроб. Агенты Чемотер. 37(4), 1993 г.
PMID: 8280208
Факторы, необходимые in vitro для удаления конъюгативного транспозона Bacteroides, CTnDOT.
Sutanto Y, DiChiara JM, Shoemaker NB, Gardner JF, Salyers AA. , Plasmid 52(2), 2004 г.
, Plasmid 52(2), 2004 г.
PMID: 15336489
Конъюгативный перенос ДНК у Streptomyces, мицелиального организма.
Тома Л., Мут Г., Плазмида 87-88(), 2016 г.
PMID: 27687731
Механизмы и барьеры горизонтального переноса генов между бактериями.
Томас К.М., Нильсен К.М., Нац. Преподобный Микробиолог. 3(9), 2005 г.
PMID: 16138099
Thomas, 2008
Характеристика различных плазмид, разлагающих 2,4-дихлорфеноксиуксусную кислоту, выделенных из почвы путем комплементации.
Top EM, Holben WE, Forney LJ., Appl. Окружающая среда. микробиол. 61(5), 1995 г.
PMID: 7646006
Усиление разложения 2,4-дихлорфеноксиуксусной кислоты (2,4-D) в почве путем распространения катаболических плазмид.
Top EM, Van Daele P, De Saeyer N, Forney LJ., Антони Ван Левенгук 73(1), 1998 г.
PMID: 9602282
Первый геномный анализ широкого круга хозяев Rhizobium sp. Штамм LPU83, представитель орегоноподобных Rhizobium sp. с низким генетическим разнообразием. группа.
Штамм LPU83, представитель орегоноподобных Rhizobium sp. с низким генетическим разнообразием. группа.
Техеризо Г.Т., Дель Папа М.Ф., Драги В., Лозано М., Джусти Мде Л., Мартини С., Салас М.Е., Сальто И., Вибберг Д., Щепановски Р., Вайднер С., Шлютер А., Лагарес А., Писторио М., J. Biotechnol . 155(1), 2011 г.
PMID: 21329739
Плазмида ризобий pLPU83a способна переключаться между различными механизмами переноса в зависимости от своего геномного фона.
Торрес Техеризо Г., Писторио М., Алтабегойти М.Дж., Сервантес Л., Вибберг Д., Шлютер А., Пулер А., Лагарес А., Ромеро Д., Бром С., FEMS Microbiol. Экол. 88(3), 2014 г.
PMID: 24646299
Конъюгативный транспозон Tn925: усиление конъюгального переноса тетрациклином у Enterococcus faecalis и мобилизация хромосомных генов у Bacillus subtilis и E. faecalis.
Torres OR, Korman RZ, Zahler SA, Dunny GM., Mol. Генерал Жене. 225(3), 1991 г.
PMID: 1850085
Конъюгативный перенос p42a из rhizobium etli CFN42, необходимый для мобилизации симбиотической плазмиды, регулируется определением кворума.
Тун-Гарридо С., Бустос П., Гонсалес В., Бром С., Дж. Бактериол. 185(5), 2003 г.
PMID: 12591886
Мобилизация горизонтально приобретенного острова 2 индуцируется в растениях фитопатогеном Pectobacterium atrosepticum SCRI1043 и включает предполагаемую релаксазу ECA0613 и определение кворума.
Ванга Б.Р., Рамакришнан П., Батлер Р.К., Тот И.К., Ронсон К.В., Джейкобс Дж.М., Питман А.Р., Окружающая среда. микробиол. 17(11), 2015
PMID: 26271942
Опины стимулируют индукцию генов vir плазмиды Agrobacterium tumefaciens Ti.
Veluthambi K, Krishnan M, Gould JH, Smith RH, Gelvin SB., J. Bacteriol. 171(7), 1989 г.
PMID: 2738020
Характеристика области ermF размером 13 тысяч оснований конъюгативного транспозона Bacteroides CTnDOT.
Whittle G, Hund BD, Shoemaker NB, Salyers AA., Appl. Окружающая среда. микробиол. 67(8), 2001 г.
PMID: 11472924
Характеристика генов, участвующих в модуляции конъюгального переноса конъюгативного транспозона Bacteroides CTnDOT.
Whittle G, Shoemaker NB, Salyers AA., J. Bacteriol. 184(14), 2002 г.
PMID: 12081954
Система конъюгации Tcp Clostridium perfringens.
Wisniewski JA, Rood JI., Plasmid 91(), 2017 г.
PMID: 28286218
TcpM: новая релаксаза, которая опосредует перенос больших конъюгативных плазмид из Clostridium perfringens.
Wisniewski JA, Traore DA, Bannam TL, Lyras D, Whisstock JC, Rood JI., Mol. микробиол. 99(5), 2015
PMID: 26560080
Интеграция и удаление CTnDOT.
Wood MM, Gardner JF., Microbiol Spectr 3(2), 2015 г.
PMID: 26104696
Интегративные и конъюгативные элементы: мозаичные мобильные генетические элементы, обеспечивающие динамический латеральный поток генов.
Возняк Р.А., Уолдор М.К., Нац. Преподобный Микробиолог. 8(8), 2010 г.
PMID: 20601965
Геномная микробная эпидемиология необходима для понимания глобальной проблемы устойчивости к антибиотикам и улучшения диагностики патогенов.
