
вакцинация от COVID-19 в столице и обязательное анкетирование в Турции – Москва 24, 15.03.2021
Самому пожилому москвичу, который вакцинировался от коронавируса, исполнилось 104 года. Более половины горожан, сделавших прививку, старше 60 лет. Всего в столице от коронавирусной инфекции привились более 700 тысяч человек. Такие цифры прозвучали во время онлайн-конференции «COVID-19. Российский и международный опыт». Специалисты отметили, что «Спутник V» показал одинаковую эффективность во всех возрастных категориях.
Прививку можно сделать, если отсутствует обострение хронических заболеваний. Среди противопоказаний есть также перенесенное вирусное заболевание в течение двух недель до вакцинации. Продолжительность действия препарата может составить более двух лет.
В Москве с января работают выездные бригады вакцинации, которые принимают горожан в торговых центрах, флагманских офисах «Мои документы» и других общественных местах.
Мировой спрос на «Спутник V» превышает предложение, однако в приоритете по-прежнему остается вакцинация внутри страны. Такую информацию озвучил Дмитрий Песков. По словам пресс-секретаря президента, Владимир Путин ежедневно получает доклады от Российского фонда прямых инвестиций. Он лично контролирует распространение вакцины за рубежом.
Между тем производство «Спутника V» может начаться в Италии и ФРГ. Об этом сообщает агентство «Рейтер», но официально эта информация пока не подтверждена. В настоящий момент Евросоюз подписал соглашения с шестью западными производителями вакцин и начал переговоры еще с двумя. Европейский медицинский регулятор одобрил четыре прививочных препарата. Однако соблюдать график поставки не получается, и некоторые страны ищут выход из ситуации самостоятельно.
Процедуру экспертизы российской вакцины специалисты ЕС начали в марте. Однако не дожидаясь официального разрешения, Венгрия и Словакия уже договорились о поставках «Спутника V». О своей заинтересованности в российской вакцине также заявили Чехия и Хорватия.
О своей заинтересованности в российской вакцине также заявили Чехия и Хорватия.
Турецкие власти утвердили новые правила въезда в страну. С 15 марта туристов пускают только после заполнения электронной анкеты, форма которой опубликована на сайте министерства здравоохранения. Ее необходимо заполнить за 72 часа до начала путешествия и предъявить при въезде в распечатанном или электронном виде. В анкету вносятся паспортные данные, контактная информация и многое другое.
После этого всем туристам автоматически присвоят код, который необходимо предъявлять по всей Турции, например при входе в торговые центры и государственные учреждения. В Стамбуле код также нужен для посещения кафе, парикмахерских, спа-салонов, театров и фитнес-центров.
Власти страны хотят при помощи кода отслеживать контакты туристов с больными коронавирусом и предупреждать их о возможном заражении. Поэтому, если анкета отсутствует или содержит некорректные данные, туристам грозят административные санкции.
Вирусные инфекции в Турции — Новости туризма
Вирусные инфекции в Турции
Казанским туристам, принимая решение о поездке в запланированный отпуск на курорты Турции, необходимо учитывать сложившуюся ситуацию с заболеваемостью вирусом Коксаки.
Данный энтеровирус распространяется контактным, водным, бытовым путём и болеют им в основном дети до шести лет. Случаи заболевания данным вирусом ежегодно фиксируются в жарких странах и составляют менее 1% от общего числа обратившихся за медицинской помощью туристов по самым разным заболеваниям.
Согласно данным представителей страховых компаний, за летний сезон (с мая по настоящий момент) этого года в общем количестве 800 тысяч застрахованных туристов зафиксированные случаи данного вида энтеровирусной инфекции составляют 83 человека, что составляет 0,1%.
Мерами профилактики при поездке с маленькими путешественниками, помимо соблюдения правил личной гигиены, в случае неблагоприятной ситуации в конкретном отеле рекомендуется ограничить посещение бассейна и детского клуба.
При появлении первых признаков заболевания у ребёнка (высокая температура, пузырьки на коже) обратиться за квалифицированной медицинской помощью. В целях профилактики при семейной поездке возможно заменить групповой трансфер из аэропорта на индивидуальный, и выяснять ситуацию с заболеваемостью в отеле.
В данный момент в Турции находится около 500 тысяч российских граждан, случаи заболевания вирусом Коксаки на фоне этого количества отдыхающих минимальны, обстановка спокойная и в большинстве отелей, ориентированных на отдых с детьми, случаев с подтверждённым медиками диагнозом не зафиксировано.
В официальных сообщениях органов исполнительной власти РФ отсутствуют решения или рекомендации по изменению или запрету полётов на курорты Турции.
В свете существующей ситуации для туристов, решивших аннулировать ранее купленный тур, вернуть средства без финансовых потерь возможно исключительно в судебном порядке (статья 14 ФЗ “Об основах туристской деятельности РФ”). Туроператор при расторжении туристом договора о приобретённом туре вправе предложить изменение сроков поездки и страны пребывания.
Будьте в курсе актуальных туристических новостей, звоните по телефону +7 (843) 567-20-20 и планируйте Ваши путешествия осенью 2021 года вместе с нами.
Хотите быть в курсе скидок и акций — Подпишитесь на нашу рассылку!
Отправляя нам свои данные, вы даете согласие на их обработку — политика конфиденциальности.
Что нужно знать о венерических заболеваниях, отправляясь в отпуск
К сожалению, иногда люди из отпуска привозят не только сувениры и яркие впечатления, но и заболевания. О некоторых из них вслух говорить неприлично. Врачи подчёркивали, что каждый год к концу лета фиксируется максимальное количество обращений пациентов с различными заболеваниями, передающимися половым путем.
О некоторых из них вслух говорить неприлично. Врачи подчёркивали, что каждый год к концу лета фиксируется максимальное количество обращений пациентов с различными заболеваниями, передающимися половым путем.
Заболевания, которые передаются половым путем, не являются характерными для каких-то конкретных стран. Хламидиоз и трихомониаз одинаково часто привозят и из Турции, и из Египта, и из Таиланда. Важно не то, где отдыхает человек, а с кем. У каждого отдыхающего по итогу могут возникнуть свои проблемы. В основном венерические заболевания привозят из отпуска люди сексуально активного возраста.
Поэтому, отправляясь в отпуск, каждый взрослый человек должен заранее подумать о последствиях приятного времяпрепровождения. Рассмотрим специфику основных венерических заболеваний, с которыми можно столкнуться сегодня.
Самыми опасными считаются вирусные инфекции, представленные Вирусом иммунодефицита человека, гепатитом, Вирусом папилломы человека, кондиломами. Вирус папилломы — это предпосылка к развитию онкологических заболеваний половых органов. Вирусной природой также обладает герпес. Если неправильно лечить или не лечить вовсе, не менее опасными считаются бактериальные инфекции: хламидиоз, сифилис, гонорея, уреаплазмоз, микоплазмоз.
Вирус папилломы — это предпосылка к развитию онкологических заболеваний половых органов. Вирусной природой также обладает герпес. Если неправильно лечить или не лечить вовсе, не менее опасными считаются бактериальные инфекции: хламидиоз, сифилис, гонорея, уреаплазмоз, микоплазмоз.
Грибковые инфекции наподобие молочницы (кандидоз) могут передаваться не только половым путем. Для их появления достаточно снижения иммунитета, гормонального сбоя или приема антибиотиков.
Как минимизировать риск заражения инфекциями, передающимися половым путём?
Вместе с кремом для загара и стандартной аптечкой, в отпуск надо брать интимные медикаменты. Речь идёт об антисептике со специальной удобной насадкой, который используют в течение 5 – 10 минут после полового акта.
Такой антисептик рекомендуется применять и женщинам, и мужчинам.
К простым способам избегания венерических заболеваний относится использование средств предохранения, отсутствие случайных половых контактов, соблюдение интимной гигиены.
Экстренная профилактика после случайных связей
- сразу после полового акта надо обильно помочиться
- с мылом вымыть руки и наружные половые органы
- произвести обработку половых органов, лобка и бёдер антисептиком
- если в течение суток не получается посетить врача, надо выпить «ударную» дозу антибиотиков как можно скорее посетить врача-уролога
Инфекционные болезни
ЧТО ВАЖНО ЗНАТЬ О ТУБЕРКУЛЕЗЕ
ТУБЕРКУЛЕЗ – инфекционное заболевание, вызываемое туберкулезной палочкой и передающееся воздушно-капельным, воздушно-пылевым путем.
Палочкой Коха иногда называют микобактерию туберкулеза (МБТ) по имени открывшего её миру немецкого микробиолога Роберта Коха. Случилось это 24 марта 1882 года. Поэтому 24 марта проводится Всемирный день борьбы с туберкулезом.
Микобактерии туберкулеза (МБТ) в окружающей среде сохраняют активность длительное время, устойчивы к воздействию многих факторов, могут подниматься периодически вверх с пылью, а значит – заражать людей.
Возможно заражение МБТ алиментарным путем. Дети могут заразиться этим путём, облизывая, пробуя «на зуб» немытые игрушки: после игры в песочнице, после падения их на асфальт. Взрослые – при использовании общей посуды. Кроме того, возможно заражение туберкулезом после употребления молока от больных туберкулезом животных. Контактный путь заражения возможен при несоблюдении правил работы с биоматериалом в лабораториях. Возможен вертикальный (внутриутробный) путь заражения – от матери к плоду во время беременности и родов.
МБТ могут поражать любые органы и ткани человека. Иммунная система человека защищает организм, в том числе от МБТ. Но при ослаблении факторов защиты возможно инфицирование и заболевание человека.
Заболеть туберкулезом может любой человек, независимо от социального положения и уровня дохода.
Инфекция, проникнув в организм любого ребёнка или взрослого, может вызвать серьезное заболевание. Поэтому очень важно проводить первичную профилактику туберкулеза новорожденным, детям, подросткам, взрослым – регулярно обследоваться.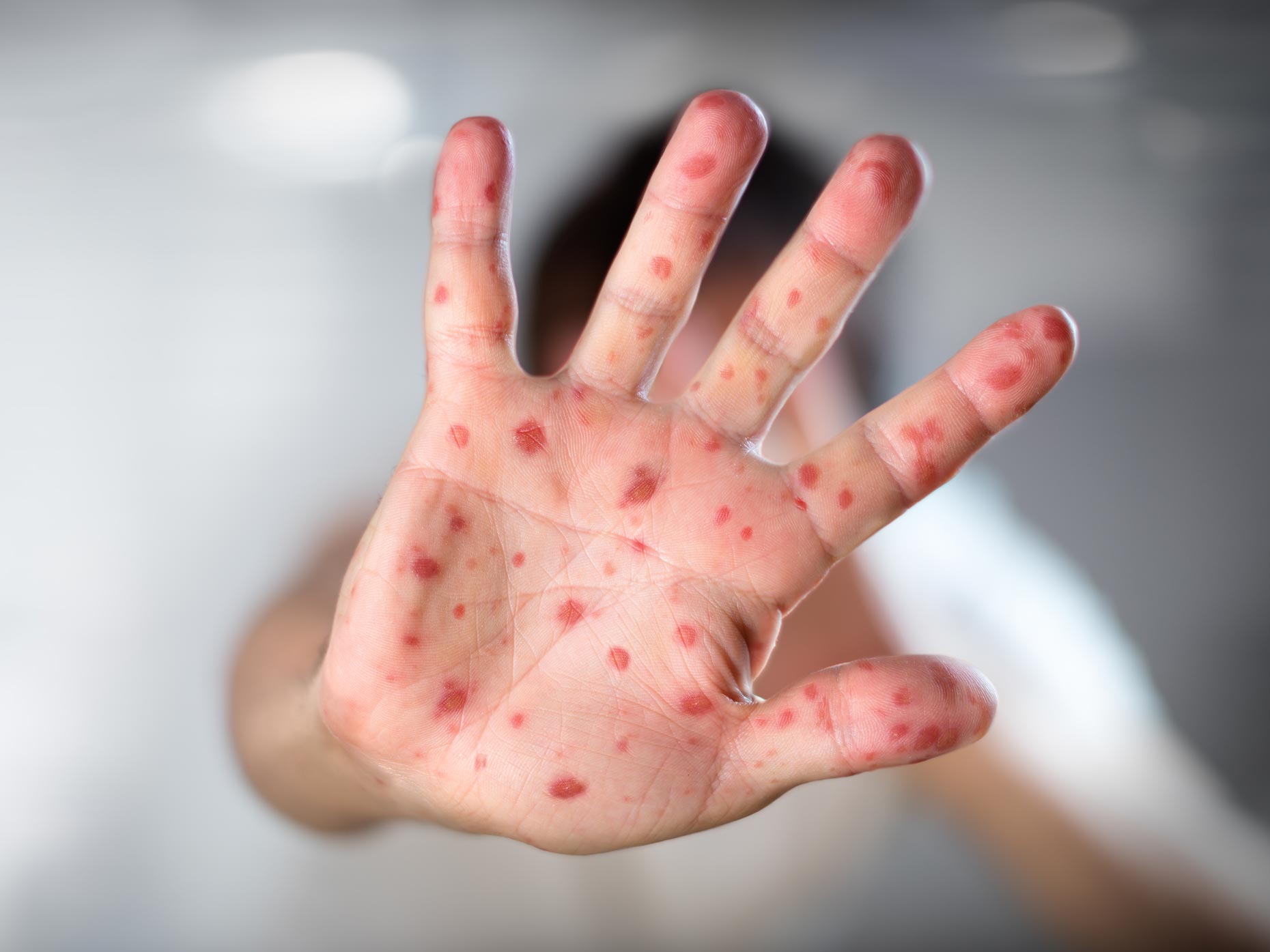
Несмотря на успехи медицины в области лечения туберкулеза заболевание легче предупредить, чем лечить.
Профилактика туберкулеза начинается с родильного дома. Национальным календарем профилактических прививок Российской Федерации предусмотрена вакцинация против туберкулеза в родильном доме. Вакцинация проводится вакциной БЦЖ-М от туберкулеза проводится в роддоме на 3-7 день жизни ребенка при отсутствии медицинских противопоказаний. Эффективность вакцинации проверена и доказана временем.
Если ребенок не был привит в роддоме, прививка проводится в поликлинике после снятия противопоказаний. До прививки и на период выработки иммунитета (2 мес.) необходимо ограничить круг взрослых, общающихся с ребенком и провести их обследование на туберкулез (флюорографию).
Дети с хроническими заболеваниями нуждаются в вакцинации больше, чем здоровые, т.к. вакцинация протекает, как правило, легко, незаметно для организма, а течение туберкулеза у таких детей, чаще всего, тяжелое и требует значительных усилий медицинских работников и самого больного на пути к выздоровлению. При решении вопроса о проведении прививки этим детям важно правильно выбрать период, обеспечивающий безопасную вакцинацию (отсутствие обострения хронического заболевания).
При решении вопроса о проведении прививки этим детям важно правильно выбрать период, обеспечивающий безопасную вакцинацию (отсутствие обострения хронического заболевания).
Уважаемые родители!
Вовремя сделайте Вашему ребенку прививку против туберкулеза!
ТУБЕРКУЛЕЗ ОПАСЕН, НО ИЗЛЕЧИМ!
Чем раньше выявлено заболевание (инфицирование), тем лучше прогноз.
Для своевременного выявления инфицирования туберкулезом используются диагностические тесты и исследования. В их числе туберкулинодиагностика (проба Манту) для детей, внутрикожная проба с аллергеном туберкулезным рекомбинантным (ДИАСКИНТЕСТ) для детей и взрослых, флюорографическое исследование для взрослых и подростков с 15-ти лет. Для выявления туберкулеза используются и другие методы диагностики, главное вовремя обратиться к врачу и не отказываться от предлагаемых исследований.
Проба Манту, туберкулинодиагностика
Туберкулинодиагностика проводится – не реже 1 раза в год.
Два раза в год проводят туберкулиновую пробу:
• Детям, не вакцинированным против туберкулеза, независимо от их возраста
• Детям, находящимся в контакте с больным туберкулезом
• Детям из групп риска по результатам туберкулинодиагностики
• Детям из групп медико-биологического риска: больным сахарным диабетом, язвенной болезнью, болезнями крови, системными заболеваниями, получающим длительно (более 1 месяца) кортикостероидную терапию, ВИЧ-инфицированным, детям с хроническими неспецифическими заболеваниями
• Детям из социальных групп риска
ЦЕЛЬ ПРОВЕДЕНИЯ ТУБЕРКУЛИНОДИАГНОСТИКИ
• Выявление больных туберкулезом
• Выявление лиц, инфицированных микобактериями туберкулеза
• Отбор детей для ревакцинации против туберкулеза.
Вопрос о возможных противопоказаниях для проведения реакции Манту должен решить врач.
Положительная туберкулиновая проба может свидетельствовать и о наличии противотуберкулезного иммунитета (поствакцинальная аллергия) и об инфицированности микобактериями туберкулеза (инфекционная аллергия).
Внутрикожная проба с аллергеном туберкулезным «ДИАСКИНТЕСТ»
Внутрикожная проба с аллергеном туберкулезным рекомбинантным, Диаскинтест» позволяет отличить туберкулезную инфекцию с высоким риском развития заболевания от реакции на вакцинацию у детей, также используется для диагностики туберкулеза у взрослых, контингентов группы риска.
Если в вашей семье ждут рождения ребенка, членам семьи из окружения беременной необходимо пройти обследование на туберкулез.
Группы риска по заболеванию туберкулезом.
- ВИЧ-инфицированные
- Пациенты, получающие терапию генно-инженерными биологическими препаратами, имунносупрессивную терапию
- Лица, попавшие в трудную жизненную ситуацию и находящиеся в организациях социальной защиты населения
- Дети, находящиеся в организациях (дома ребенка, интернаты, приюты и др.)
- Лица, попавшие в трудную жизненную ситуацию, при обращении в медицинские организации за медицинской помощью
- Лица, находящиеся в медицинских организациях психиатрического профиля
- Пациенты, состоящие на учете в филиалах Государственного казенного учреждения здравоохранения города Москвы “Московский научно-практический центр наркологии
- Освобожденные из следственных изоляторов и исправительных учреждений Федеральной службы исполнения наказаний России
- Декретированные контингенты при профилактических медицинских, в т.
 ч. медицинские работники
ч. медицинские работники - Больные с хроническими соматическими заболеваниями, в т.ч. сахарным диабетом.
Когда можно заподозрить у себя туберкулез?
Туберкулез часто протекает бессимптомно. Его можно выявить только при специальном обследовании. Для этого существует диспансеризация. Однако у некоторых пациентов могут быть клинические проявления заболевания.
Заподозрить туберкулез и сдать мокроту для анализа на МБТ необходимо, если у Вас или у Вашего ребёнка есть один или несколько из этих симптомов:
- кашель более двух-трёх недель
- боль в грудной клетке, связанная с дыханием
- длительное повышение температуры
- потеря веса, не связанная со специальным похуданием
- слабость, усталость и быстрая утомляемость
- повышенная потливость, часто – ночная потливость
- прожилки крови в мокроте
Если один или несколько из вышеперечисленных симптомов есть у Вас или Вашего ребёнка,
- обратитесь к врачу-терапевту, врачу-педиатру.
- пройдите обследование на туберкулез, чтобы исключить эту инфекцию или выявить заболевание как можно раньше и быстрее вылечиться!
Как не заболеть туберкулезом?
• Обязательно делайте новорожденному прививку БЦЖ-М от туберкулеза.
• Соблюдайте правила личной гигиены.
• Мойте руки перед едой, после контакта с пылью и любыми грязными предметами, после нахождения в общественных местах (общественном транспорте, магазине, поликлинике и больнице).
• Не трогайте грязными руками глаза и лицо.
• Не позволяйте ребёнку брать в рот немытые игрушки (упавшие на улице или те, с которыми он играл на улице, в детском саду, в песочнице).
• Не берите на прогулку малышу мягкие игрушки.
• Избегайте контактов (не находитесь вблизи) с кашляющими людьми, особенно – с длительно кашляющими.
• Чаще бывайте на улице: гуляйте в солнечную погоду.
• Старайтесь не переохлаждаться и соблюдать режим дня.
• Следите за питанием. Питание должно быть рациональным и сбалансированным, содержать овощи, фрукты.
В случае если в Вашем окружении кто-то заболел туберкулезом, обязательно пройдите профилактическое обследование (флюорографию, общий анализ крови, выполните кожные тесты: пробу Манту с 2 ТЕ, Диаскинтест).
Если в Вашем доме или в квартире кто-то заболел туберкулезом,
необходимо обязательно:
• Пройти обследование на туберкулез (детям и подросткам – пробу Манту, взрослым и подросткам – флюорографию, Диаскинтест и др.).
• Не игнорируйте приглашение на обследование!
• Соблюдайте назначения и рекомендации лечащего врача, в т.ч. по соблюдению санитарно-гигиенического режима.
Осторожно, коронавирусная инфекция! — Объявления
По официальным данным в настоящее время зарегистрировано в 25 странах мира более 1200 случаев инфицирования человека ближневосточным респираторным синдромом коронавируса (БВРС-КоВ), из них 498 случаев — с летальным исходом. Большинство случаев зарегистрировано в странах Ближнего Востока, а так же в США, Австрии, Германии, Франции, Греции, Италии, Нидерландах, Турции, Соединённом Королевстве, Южной Корее.
Коронавирусная инфекция — острое вирусное заболевание с преимущественным поражением верхних дыхательных путей, чаще в виде ренита, или ЖКТ по типу гастроэнтерита. Возбудитель — РНК-содержащий вирус. Патогенными для человека признаны респираторные и кишечные коронавирусы. У человека доминируют респираторные коронавирусные инфекции, регистрируемые повсеместно как «банальная простуда» или ОРВИ.
Резервуар и источник инфекции — больной человек, период контагиозности больного неопределённо долгий. К вирусу чувствительны все возрастные группы населения, однако преимущественно болеют дети и подростки. Входными воротами для возбудителя может стать слизистая оболочка верхних дыхательных путей; в этом случае болезнь развивается по типу ОРВИ с тяжелым респираторным синдромом (атипичная пневмония), может присоединиться почечная недостаточность. При реже встречающемся фекально-оральном механизме передачи заболевание проявляется в виде гастроэнтерита. Заболевание сопровождается синтезом антител, не защищающих от повторного заражения вирусом других серологических типов.
В целях предупреждения заражения коронавирусной инфекцией гражданам настоятельно рекомендуется воздержаться от поездок в страны Ближнего Востока (Саудовская Аравия, Катар, ОАЭ) и Южную Корею без острой необходимости. При выезде в указанные страны в случае необходимости рекомендуется использовать защитные маски и воздержаться от посещения мест массового скопления людей и контактов с больными людьми с высокой температурой, а также избегать контакта с больными животными, не употреблять в пищу мясо, не прошедшее надлежащеё тепловой обработки, регулярно проветривать помещение, делать влажную уборку и соблюдать правила личной гигиены.
Филиал ФБУЗ «ЦГиЭ в РО» в Аксайском районе
Простуда или Коксаки? Страховщики рассказали, чем чаще болеют дети на отдыхе
Самой распространенной причиной обращения за помощью на отдыхе является совсем не вирус Коксаки, а другие болезни
«Бушующий» вирус Коксаки продолжают обсуждать в соцсетях. Общественность разделилась на два лагеря – одни утверждают, что их клиенты чудом спаслись от ужасной болезни, другие пишут, что это полный бред и раздутая история. Давайте еще раз рассмотрим ситуацию, теперь с точки зрения страховой статистики.
В Турции Коксаки – нет!
Глава Федерации отельеров Турции Осман Айык рассказал журналистам, что вирус Коксаки в страну завезли сами российские туристы. По его словам, болезнь действительно была распространена на территории курортов, однако на данный момент у турецких детей она не наблюдается.
«Турция решила проблему. Инкубационный период этого заболевания составляет две недели. В Анталье русские отдыхают в среднем семь-восемь дней. Если рассуждать логически, они могут приезжать уже с этим заболеванием. Два года назад были частые жалобы, но в этом году мы не получили ни одной. До сих пор из наших отелей не поступило ни одного сообщения в этом ключе. Турпоток из России невероятный. Каждый раз, когда так бывает, в российской прессе обязательно выходят такого рода недостоверные новости против нас», – рассказал Айык турецкой Hurriyet.
Впрочем, не следует воспринимать эти слова буквально. Надо сделать поправку на то, что выступает лицо заинтересованное, кроме того, возможны неточности при переводе с турецкого. Давайте посмотрим на данные страховых компаний.
Юлия Алчеева, исполнительный директор компании ERV, сообщает, что в начале сезона (май, июнь) был зарегистрирован только 81 случай заболевания Коксаки в Турции, это порядка 4 % от всех обращений туристов. Причем в июле ситуация начала улучшаться, и в целом эпидемиологический порог не превышен.
А в «АльфаСтраховании» рассказали, что беспокойство общественности преувеличено. «Мы не видим аномального всплеска обращений по инфекционным заболеваниям на территории Турции на начало августа», – прокомментировал HotLine.travel Юрий Нехайчук, официальный представитель компании. Беспокоиться нужно совсем о другом.
Чем болеют дети на отдыхе
По статистике «АльфаСтрахования», которой компания поделилась в пресс-релизе от 20 июня, в рейтинге причин для обращения за медпомощью тех самых «Коксак» в принципе нет. Более 30 % страховых случаев среди юных туристов приходится на воспалительные заболевания дыхательных путей (фарингит, тонзиллит) и отиты. От них чаще всего страдают дети в возрасте четырех – семи лет. На втором месте острые респираторные инфекции (18,2 %), на третьем – энтериты и гастриты (14,38 %). Обе проблемы чаще всего встречаются у малышей до трех лет. На четвертой позиции рейтинга травмы (7,41 %). Замыкает топ-5 самых частых «детских» заболеваний на отдыхе конъюнктивит (1,81 %). Этот рейтинг касается не только Турции, но и других стран, в которых отдыхают россияне с детьми.
«Стрессовая смена обстановки в путешествии, акклиматизация, перепады температур на солнце и в тени, обилие охлаждающих напитков, попавшая во время купания в уши вода – всё это может привести к воспалительным процессам и стать причиной болезни ребенка», – рассказал Антон Колегов, главный андеррайтер управления страхования путешествующих компании «АльфаСтрахование».
Идем дальше. В рейтинге стран, составленном компанией по объему страховых случаев с маленькими туристами, лидирует Болгария (30,25 %), второе место за Испанией (12,89 %), замыкает тройку стран Таиланд (6,8 %). Турция лишь на седьмом месте с 3,46 % обращений. Это данные на начало июня, которые не охватывают «высокий» сезон, так что соотношение теоретически может измениться. Тем не менее вряд ли картина поменяется кардинально.
Вот и научные исследования показывают, что настоящая опасность исходит с другой стороны. «Газета.ru» в своей недавней публикации утверждает, что в летние месяцы бактериальные заболевания распространены гораздо сильнее, чем вирусные, в том числе Коксаки.
Реальная угроза, исходящая от бактерий летом, – пищевые отравления. Высокая температура и влажность создают благоприятную среду для размножения бактерий. Служба контроля безопасности пищевых продуктов США предупреждает, что если при температуре воздуха выше +32 °C еда более часа пролежала вне холодильника, лучше ее не есть.
Рекомендации, которые дают специалисты службы для предотвращения отравлений, просты: тщательно мыть руки после посещения туалета, смены подгузников ребенку и контакта с животными, хранить приготовленную пищу отдельно от сырых продуктов и тщательно прожаривать мясо.
Если на первом месте среди виновников страховых случаев у детей стоят болезни дыхательных путей и воспалительные заболевания, почему никто не поднимает панику по поводу эпидемии отита в Анталье? Видимо, слово Коксаки звучит экзотично, малопонятно и потому загадочно. А люди боятся того, чего не понимают.
Поставить точку в вопросе «Стоит ли бояться Коксаки в Турции летом-2017?» можно следующим комментарием под статьей на HotLine.travel 29 июля: «У нас сейчас все признаки вируса Коксаки, и заболели мы не в Турции, а в РОСТОВЕ-НА-ДОНУ».
Так что посоветуйте туристам соблюдать правила гигиены, не оставлять детское питание под палящими лучами солнца, побольше купаться в море, а не в бассейне – и тогда, скорее всего, этой профилактики вируса Коксаки будет достаточно, чтобы про него не вспоминать.
Представительство в Турции | Клиника МЕДЕЛ
ПредставительствоМногопрофильной Клиники МЕДЕЛ в Турции
GABDELGANI NURULLİN
Габдельгани Нуруллин
Официальный
представитель
Клиники МЕДЕЛ в Турции
Tel: +9 050 763-77-775
Watsap: +7 937 777-55-75
Одна из востребованных услуг —
ЭкстраКорпоральное ОплодотворениеЭКО (от лат. extra — сверх, вне и лат. corpus — тело, то есть оплодотворение вне тела, сокр. ЭКО́) — вспомогательная репродуктивная технология, чаще всего используемая в случае бесплодия.
Где сделать искусственное оплодотворение (ЭКО)?
Важно сделать правильный выбор при принятии решения, в какой клинике вы будете получать лечение искусственного оплодотворения. На пути к тому, чтобы стать родителем, вы сталкиваетесь с проблемами фертильности, не только с медицинской, но и эмоциональной и финансовой точек зрения.
Поэтому мы рекомендуем надежного партнера Клиники МЕДЕЛ в Турции.
Эффективность процедурыТурецкие клиники по ЭКО осуществляют порядка 50 000 процедур искусственного оплодотворения за год, немалая часть пациентов приезжает из-за границы. Востребованность услуг турецких репродуктологов объясняется высокой эффективностью: прядка 50% успеха у молодых пар и около 40% — у женщин, чей возраст перешел отметку в сорок лет.
Преимущества турецких клиникВ клиниках Турции используют наиболее современные диагностические методики, которые позволяют точно определить причину мужского и женского бесплодия, и выбрать наиболее перспективный вариант проведения ЭКО. Турецкие медики активно сотрудничают со своими коллегами из США, Германии, Израиля, это дает возможность перенимать передовой опыт и успешно внедрять его в свою практику. К проблеме бесплодия тут подходят комплексно: в выработке стратегии проведения ЭКО привлекаются гинекологи, андрологи, репродуктологи, эндокринологи и другие узкие специалисты.
Цены на ЭКО в ТурцииСтоимость зависит от выбранного метода, состояния здоровья пациентов и других индивидуальных факторов. Цена на ЭКО в турецких клиниках колеблется от 2500 до 5000 евро.
Высокая эффективность лечения и умеренная стоимость ЭКО в Турции – главные факторы, которые побуждают пациентов выбирать для проведения процедуры именно эту страну. Отзывы пациентов также отмечают благожелательное отношение медперсонала, комфортные условия пребывания в клиниках, возможность заказать услуги переводчика и качественный.
ЭКО по программе от партнера Клиники МЕДЕЛКлиника в г. Анталия: GELECEK TUP BEBEK MERKEZI ANTALYA.
Услуги, входящие в стоимость пакета ЭКО от 2500 евро:1. Осмотр пациентов;
2. Спермограмма;
3. Гепатит-тест (An HCV-An HBS-HbsAG-An HiV;
4. Процедура ЭКО;
5. Медикаменты для процедуры ЭКО;
6. Медикаменты после процедуры ЭКО;
7. Трансфер по городу (аэропорт, место размещения и Клиника).
Следующий шаг
После того, как вы сделали правильный выбор, следующим шагом является связаться с нашим партнером для первоначальной консультации.
Доктор Мете Ишикоглу
Специалист по гинекологии и акушерству
Специалист по эндоскопии
Образование и Научные степени
1. Университет Бакалавр медицинского факультета Военно-медицинской академии Гюльхане Кандидат наук / Профессиональное образование / Специализация в области медицины Акушерство и гинекология.
2. Стамбульский университет Медицинский факультет Стамбула.
Членство в научных организациях:
1. Турецкое общество гинекологии и акушерства
2. Ассоциация репродуктивной эндокринологии по бесплодию и методам вспомогательной репродукции (TSRM)
3. Турецкое общество урогинекологов
4. Общество гинекологической эндоскопии (JED)
5. Европейское общество репродуктивной медицины и эмбриологии (ESHRE)
6. Международное общество предимплантационной генетической диагностики (PGDIS)
7. Всемирная ассоциация репродуктивной медицины (WARM)
Кишечные вирусы энтерита индейки
Вирусная болезнь. 2014 июн; 25 (2): 173–185.
, , иНареш Джиндал
Департамент ветеринарного здравоохранения и эпидемиологии, Колледж ветеринарных наук, Университет ветеринарии и зоотехники Лала Ладжпат Рай, Хисар, 125 004 Индия
9004, Индия. К. МорКафедра ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Св.Paul, MN 55108 USA
Sagar M. Goyal
Департамент ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Сент-Пол, Миннесота 55108 США
Департамент ветеринарного здравоохранения и эпидемиологии, Колледж Ветеринарные науки, Университет ветеринарии и зоотехники Лала Ладжпат Рай, Хисар, 125 004 Индия
Департамент ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Св.Paul, MN 55108 USA
Автор, ответственный за переписку.Поступило 2 октября 2013 г .; Принято 27 января 2014 г.
Эта статья цитируется в других статьях в PMC.Abstract
Здоровье кишечника очень важно для получения максимальной отдачи с точки зрения увеличения веса и яйценоскости. Кишечные заболевания, такие как комплекс энтерита домашней птицы (PEC) у индеек, не позволяют максимально реализовать их производственный потенциал. Был вовлечен ряд вирусов, бактерий и простейших, но первичная этиология окончательно не установлена.Раньше электронная микроскопия использовалась для обнаружения кишечных вирусов, которые определялись исключительно на основе их морфологии. С появлением методов быстрой молекулярной диагностики и секвенирования нуклеиновых кислот нового поколения исследователи добились больших успехов в идентификации и характеристике вирусов, связанных с PEC. Молекулярные методы также помогли нам идентифицировать патогены, которые ранее не были известны. Региональные и национальные исследования выявили наличие нескольких различных кишечных вирусов в PEC, включая ротавирус, астровирус, реовирус и коронавирус, по отдельности или в комбинации.По-прежнему могут существовать неизвестные патогены, которые могут прямо или косвенно играть роль в энтерите у индеек. Этот обзор будет посвящен роли коронавируса, ротавируса, реовируса и астровируса индейки в энтерите индейки.
Ключевые слова: Энтерит, Индейки, Коронавирус, Ротавирус, Реовирус, Астровирус
Введение
На протяжении многих лет у индеек было описано несколько кишечных заболеваний, а именно: коронавирусный энтерит индеек, синдром мальдигестии, синдром ранения и задержки роста у индеек, синдром мальабсорбции у домашней птицы, синдром энтерита и смертности домашних птиц (PEMS), пиковая смертность индеек и вирусный энтерит индейки.Все эти синдромы были включены в комплекс энтерита домашней птицы (PEC), общий термин, обозначающий инфекционные кишечные заболевания молодняка индеек [6]. ПЭК — это экономически важное заболевание молодняка индеек, характеризующееся энтеритом, диареей, умеренной или выраженной депрессией роста, нарушением использования корма, плохой прибавкой веса и в некоторых случаях высокой смертностью, как при ПЕМС [6]. Ряд вирусов (коронавирус, калицивирус, реовирус, астровирус, ротавирус, пикорнавирус, пикобирнавирус, парвовирус и аденовирус), бактерий ( Escherichia coli , Salmonella spp., Clostridia, Campylobacter и Enterococcus ) и простейшие (кокцидии и криптоспоридиумы) участвуют в PEC [6, 18, 47, 73, 105, 109]. Недавно у индюшат Миннесоты были описаны два новых синдрома, а именно «синдром энтерита домашней птицы», или PES, и «синдром легкой индейки», или LTS. ПЭС — это инфекционное кишечное заболевание молодых индеек в возрасте от 1 дня до 7 недель, которое характеризуется диареей, депрессией и летаргией с бледным кишечником и / или чрезмерно жидким содержимым слепой кишки [47].LTS — это проблема индеек товарного возраста, имеющих меньшую массу тела по сравнению с их стандартными породными характеристиками [68]. Мы полагаем, что энтерит у домашних птиц в молодом возрасте создает условия для будущего развития LTS у более старых птиц. Кроме того, мы рассматриваем как PES, так и LTS как часть PEC.
Кишечные вирусы являются частой причиной первичного повреждения желудочно-кишечного тракта (ЖКТ) молодых цыплят, создавая благоприятную среду для роста бактерий и / или простейших и вызывая дальнейшее повреждение кишечника.Возможно, что повреждение кишечника сначала кишечными вирусами, а затем вторичными патогенами может привести к необратимым изменениям в организме хозяина, что может привести к снижению массы тела при продаже. Цель этого обзора — подробно обсудить четыре наиболее важных вируса, вызывающих энтерит индейки, а именно коронавирус, реовирус, ротавирус и астровирус.
Коронавирус
Коронавирусный энтерит индейки — острое заразное заболевание, характеризующееся тупостью, депрессией, диареей и сниженным набором массы тела.Заболевание было впервые описано в 1940-х годах в штате Вашингтон, США. Возбудитель не был идентифицирован, и заболевание было названо «грязевой лихорадкой». Однако после выявления в этих случаях коронавируса индейки (TCoV) название было изменено на «коронавирусный энтерит индеек». В 1970-х годах эта болезнь имела самые разрушительные последствия для индеек из Миннесоты. Были предприняты шаги по искоренению болезни, которые привели к элиминации этого вируса из Миннесоты. В настоящее время TCoV спорадически регистрируется из регионов Северной Америки и других стран, где выращивают индейку.TCoV также связан с PEMS, острым высококонтагиозным кишечным заболеванием, характеризующимся депрессией, анорексией, диареей и высокой смертностью индеек.
Геном
Коронавирусы представляют собой плеоморфные частицы с оболочкой, примерно сферической формы, диаметром от 50 до 200 нм. Согласно последнему предложению Международного комитета по таксономии вирусов (ICTV), семейство Coronaviridae включает два подсемейства: Coronavirinae и Torovirinae. Coronavirinae состоит из трех родов: Alphacoronavirus , Betacoronavirus и Gammacoronavirus [54], из которых последний включает коронавирусы птиц. Геном коронавируса состоит из одноцепочечной РНК с положительным смыслом. Полноразмерный геном состоит из 27 632 нуклеотидов плюс 3 ‘поли (А) хвост. Есть две открытые рамки считывания (ORF), ORF 1a и 1b, которые находятся в первых двух третях генома с девятью дополнительными ORF ниже по течению.Область между генами белков мембраны (M) и нуклеокапсида (N) содержит три потенциальных малых ORF: (1) ORF-X, (2) ранее не охарактеризованную ORF со связанной предполагаемой TRS в гене M (очевидно, общей для всей группы III coronaviruses) и (3) ранее описанные ORF 5a и 5b. TCoV не содержит гена гемагглютинин-эстеразы [29].
Сообщалось о вариабельности в различных генных сегментах TCoV. Сравнение последовательностей полной 3′-концевой области, кодирующей структурный белок четырех TCoV, с соответствующими последовательностями вируса инфекционного бронхита (IBV) показало, что TCoV, вероятно, имеет то же происхождение, что и IBV, и приобретенные последовательности S-гена для тропизма кишечника индейки во время процесс эволюции [61].Lin et al. [60] сообщили, что последовательность S-белка TCoV имеет только 33,8–33,9% идентичности с IBV, хотя оба вируса сгруппированы в одну и ту же геномную линию. Полноразмерные последовательности гена spike (S) французского происхождения TCoV обладали 98% идентичностью как на уровне нуклеотидов, так и на уровне аминокислот [65]. Однако эти вирусы показали 65 и 60% сходства с североамериканскими TCoV и 50 и 37% сходства с IBV на уровне нуклеотидов и аминокислот, соответственно.
Заболеваемость и распространение
В отличие от обычно неопределенных синдромов вирусных кишечных заболеваний, TCoV-энтерит можно напрямую отнести к инфекции, вызванной TCoV [35, 38, 41].TCoV распространен во всем мире и был обнаружен у пораженных энтеритом индеек из Северной Америки, Южной Америки и Европы [34, 59, 65, 70, 98]. Было обнаружено, что 37% кишечных проб от больных стад индейки в Европе оказались положительными на TCoV [65]. В недавнем исследовании, проведенном в Бразилии, большинство образцов кишечника, взятых как у пораженных энтеритом, так и у здоровых индеек, оказались положительными на TCoV [70]. В этом исследовании 55,3% TCoV-положительных образцов также были положительными на другие кишечные вирусы.Недавно были зарегистрированы вспышки коронавирусного энтерита из Арканзаса и Северной Каролины в США с тяжелой диареей и смертностью [19, 83, 100, 106], что указывает на важность постоянного эпидемиологического надзора за этим вирусом. TCoV, по-видимому, отсутствует в синдромах ПЭК без ПЭМС [44, 45, 62, 68, 75, 105]. Например, коронавирус не был обнаружен, когда в Калифорнии были протестированы 2400 образцов от домашних птиц с клиническими признаками энтерита [105]. Аналогичным образом, в исследовании 33 стай индеек, пораженных энтеритом, в США, Pantin-Jackwood et al.[75] не нашли TCoV. Мы также не обнаружили TCoV в случаях PES и LTS [44, 45, 68].
TCoV может поражать птиц любой возрастной группы, но вызывает высокую смертность молодых индюшат в возрасте 1–4 недель. Смертность колеблется от менее 10 до 50%. Индюки, по-видимому, являются единственным естественным хозяином TCoV; при экспериментальной инокуляции TCoV вызывал заболевание у домашних птиц, но не у цыплят [42]. Репликация TCoV происходит в энтероцитах, выстилающих апикальную часть ворсинок кишечника. Разрушение эпителия ворсинок кишечника приводит к диарее, нарушению пищеварения и мальабсорбции, что может привести к снижению прибавки в весе у индюшат.
Передача
Вирус присутствует в основном в кишечнике и фекалиях пораженных домашних птиц, следовательно, передача вируса осуществляется фекальным оральным путем. Прямая передача происходит при прямом контакте с инфицированными птицами, тогда как косвенная передача происходит механическим путем через рабочих и / или зараженное оборудование. Однако в экспериментальном исследовании антиген TCoV также может быть обнаружен в придаточных пазухах носа и слезной добавочной железе (железе Хардера) инфицированных цыплят в течение 14 дней после инокуляции [33].Вирус выделяется с фекалиями индеек в течение многих недель после выздоровления от клинического заболевания. Большинство случаев заражения происходит летом (май – август), иногда осенью. TCoV может сохраняться в течение длительного периода времени и может легко передаваться между птицефабриками в прохладной окружающей среде.
Клинические признаки и патология
Индюки, пораженные TCoV, сбиваются в кучу и демонстрируют взъерошенные перья, снижение потребления корма и воды, влажный помет и потерю живой массы.У пожилых птиц может наблюдаться депрессия, диарея и задержка роста [34]. Помет водянистый и пенистый, может содержать слизь, от зеленого до коричневого цвета. Клинические признаки могут сохраняться до 2 недель, а восстановление веса, если оно будет достигнуто, может занять несколько недель. У несушек наблюдается снижение яйценоскости [3] из-за аномальной пигментации яиц. Птицы в возрасте от 1 до 4 недель, у которых наблюдаются диарея, обезвоживание и задержка роста на 40% или более, считаются страдающими PEMS. Существует две формы PEMS: (1) тяжелая форма i.е. рост нравственности индеек (SMT), который определяется как смертность, равная или превышающая 1% в течение 3 дней подряд или 9% в возрасте от 8 до 28 дней; и (2) легкая форма, которая определяется как смертность более 2%, но менее 9% в течение того же периода времени, но не равная или не превышающая 1% в течение трех дней подряд. В экспериментальном исследовании, как заболеваемость, так и смертность регистрировались у домашних птиц, экспериментально инокулированных TCoV в возрасте 2 дней, однако птицы, инфицированные в возрасте 28 дней, имели значительную задержку роста, но не демонстрировали смертности [32].
TCoV может действовать синергетически с другими патогенами, увеличивая тяжесть заболевания, или может предрасполагать птиц к вторичным инфекциям. У цыплят, инокулированных только TCoV, развилась умеренная депрессия роста без гибели, а у свиней, инокулированных только энтеропатогенной Escherichia coli (EPEC) , клиническое заболевание не развилось. Однако инокуляция цыплят TCoV и EPEC одновременно вызвала повышение смертности, подавление роста и прикрепление / стирание повреждений [38].
У пораженных птиц наблюдается задержка роста и обезвоживание. Кишечник заметно увеличен и заполнен рыхлым желтым содержимым. Стенки кишечника вялые и бледные. Caeca также вздуты с пенистым и водянистым содержимым. Крупные изменения, такие как атрофия поджелудочной железы, селезенки и бурсы Фабрициуса, также могут наблюдаться наряду с расширением желчного пузыря и мочеточников. У инфицированных кур наблюдаются более выраженные поражения яичников с деформированными, плохо развитыми яичниками и скоплением казеозного экссудата в брюшине.Также наблюдаются спайки брюшины яйцевода.
В зависимости от тяжести инфекции наблюдается от легкого мультифокального до тяжелого энтерита с атрофией ворсинок. Наблюдается слущивание эпителиальных клеток с застоем и обширной инфильтрацией лимфоцитов и гетерофилов в собственную пластинку ворсинок. Кроме того, также может наблюдаться истощение фолликулярных лимфоидов и атрофия коркового вещества тимуса.
Диагноз
Клинические признаки, такие как диарея, депрессия, скученность и задержка роста, указывают на кишечную инфекцию.Высокая смертность молодняка может указывать на ПЕМС.
Выделение вируса, электронная микроскопия (ЭМ), обнаружение вирусной РНК и серология могут помочь в диагностике. Вирус можно выращивать в зародышах индейки или куриных яйцах, но его трудно выращивать в клеточных культурах. ЭМ с отрицательным контрастом может выявить вирусные частицы размером ~ 50–200 нм с типичной морфологией в форме короны в содержимом кишечника. Прямые и непрямые флуоресцентные тесты на антитела и ELISA использовались для обнаружения антигена TCoV и антител соответственно [30, 31, 37].О высокой распространенности TCoV сообщалось от заводчиков (71,1%) и мясных индеек (56,7%) в Онтарио, Канада [31]. В Арканзасе 64,2% образцов сыворотки дали положительный результат на антитела [30].
Доступны молекулярные тесты для обнаружения и дифференциации TCoV от других кишечных вирусов. Доступны различные тесты полимеразной цепной реакции с обратной транскрипцией (RT-PCR), нацеленные на 3′-нетранслируемую область (UTR), S, N и / или M ген TCoV [11, 12, 16, 89, 92, 104]. Также была разработана ОТ-ПЦР в реальном времени, которая является наиболее чувствительным и специфическим тестом, с помощью которого можно количественно определить вирусную нагрузку в тканях индейки, а также в образцах фекалий [15].Прямая иммуногистохимия была рекомендована в качестве альтернативы ОТ-ПЦР для обнаружения TCoV в полевых образцах [14].
Профилактика и контроль
Специального лечения или вакцины нет. Биобезопасность, при которой особое внимание уделяется управлению подстилкой и надлежащей утилизации мертвых птиц и использованного подстилки, похоже, единственный способ предотвратить распространение инфекции. Кроме того, общие меры, такие как уменьшение влажности подстилки, использование антибиотиков для борьбы с вторичными бактериальными инфекциями и улучшение гигиены, могут минимизировать последствия энтерита TCoV.
Реовирус
Птичий реовирус был впервые выделен от кур, страдающих хроническим респираторным заболеванием, и получил название возбудитель Фейи-Кроули (FC). Позже возбудитель FC был охарактеризован как реовирус. Однако у кур птичий реовирус вызывает множество синдромов, включая вирусный артрит / теносиновит. Недавно реовирусы были выделены от артрита / тендосиновита индейки и получили название TARV (реовирус артрита индейки) [69]. Реовирусы в основном бывают двух типов: фузогенные и нефузогенные.Слитые вирусы обладают способностью вызывать слияние инфицированных клеток, что приводит к образованию многоядерных синцитий, в то время как нефузогенные вирусы не вызывают слияния инфицированных клеток. К нефузогенным типам относятся, в основном, реовирусы млекопитающих (MRV), тогда как слитные реовирусы поражают млекопитающих, птиц и рептилий.
Геном
Реовирус принадлежит к роду Orthoreovirus в семействе Orthoreoviridae. Вирус без оболочки, имеет икосаэдрическую симметрию и размер частиц 70–80 нм.Он содержит геном двухцепочечной (ds) РНК из десяти сегментов, которые делятся на три класса, называемые большими (L), средними (M) и малыми (S), в зависимости от их характера миграции при электрофорезе в полиакриламидном геле. Гены L и M далее подразделяются на три сегмента каждый (L1, L2, L3 и M1, M2, M3, соответственно), в то время как ген S имеет четыре сегмента (S1, S2, S3, S4) [7].
Геном реовируса имеет 12 открытых рамок считывания, которые кодируют восемь структурных и четыре неструктурных белка.Структурные белки являются важной частью вирионов потомства, в то время как неструктурные белки не присутствуют в зрелом вирионе и экспрессируются только в инфицированных клетках. Структурными белками, кодируемыми генами L, M и S, являются лямбда (λ), мю (μ) и сигма (σ) соответственно. Три структурных белка λA, λB и λC кодируются сегментами L гена L1, L2 и L3 соответственно. Сегменты M1 и M2 кодируют два структурных белка (μA и μB) соответственно, в то время как сегмент M3 кодирует неструктурный белок (μNS).Три σ-белка σC, σA и σB кодируются сегментами S1, S2 и S3 соответственно, тогда как сегмент S4 кодирует неструктурный белок σNS. Сегмент S1 кодирует два дополнительных неструктурных белка, а именно p10 и p17 [7, 10].
На основании анализа последовательностей различных генов сообщалось, что реовирусы индейки отличаются от реовирусов кур [20, 44, 45, 52, 88]. Например, сходство нуклеотидной и аминокислотной последовательности сегмента S3, кодирующего сигма-B-белок реовируса индейки (TRV), было обнаружено примерно 61 и 78-80%, соответственно, по сравнению с изолятами куриного реовируса [88].Day et al. [20] сообщили, что белок сигма C гена S1 TRV имеет 57% аминокислотную идентичность с таковой у куриного реовируса (эталонный штамм S1133), тогда как белки p10 и p17 имеют 72 и 61% идентичности, соответственно, с соответствующими белками S1133.
Белок сигма С реовируса млекопитающих и слитый реовирус Нельсона Бэй на 25–28% имеют аминокислотную идентичность с белком сигма С TRV. Ранее мы сообщали, что сходство последовательностей гена S4 на уровне нуклеотидов TRVs было 71.1–74,2% по сравнению с реовирусами куриного происхождения [45]. Эти исследования показывают, что TRV образуют отдельную группу по сравнению с куриными и другими изолятами птичьего реовируса. В результате TRV потенциально можно рассматривать как отдельный вид вируса в подгруппе 2 рода Orthoreovirus . Эта точка зрения дополнительно подтверждается экспериментальными исследованиями, в которых TRV не смогли вызвать признаки заболевания у цыплят, хотя они могут реплицироваться на низких уровнях в кишечном тракте цыплят.Более того, высокопатогенный штамм реовируса куриного происхождения (ARV-1733), вызывающий высокую смертность цыплят, не вызывал клинических признаков у промышленных индеек и имел лишь легкие клинические признаки у индюков, свободных от специфических патогенов [93].
Заболеваемость и распространение
Около 85–90% изолятов реовируса не являются патогенными. Патогенные штаммы реовирусов были связаны с различными заболеваниями кур и индеек. У индеек этот вирус был обнаружен при нескольких различных кишечных заболеваниях, а именно LTS, PEC, PES, миокардите, а недавно и при артрите / теносиновите [6, 28, 39, 44, 47, 68, 69, 90, 91].При исследовании 33 стад индюшат 46% стад оказались положительными на реовирусы [75]. Jindal et al. [44] проверили содержимое кишечника 43 стад, пораженных PES, в 2007–2008 гг. И сообщили о наличии реовируса в 40% стад. Энтерит, вызванный реовирусом, в основном встречается у молодняка индейки в возрасте 1–7 недель, а заболеваемость выше у птиц в возрасте 1–3 недель, которая уменьшается с увеличением возраста.
Реовирусы обнаружены также у практически здоровых кур и индеек [45, 68, 75].При исследовании пяти здоровых родительских стад индюшат 10,4% из 193 протестированных образцов (в возрасте от 1 до 9 недель) были положительными на реовирус [45]. Тестирование образцов кишечника 33 промышленных стад индейки из всех регионов США в течение 2005 и 2006 гг. Выявило присутствие реовируса в 45,5% стад [75].
Передача
Горизонтальная передача фекально-оральным путем является обычным явлением. Из-за стабильности вируса в окружающей среде также может происходить механическая передача.Различные штаммы реовируса различаются по способности распространяться по горизонтали. Подозревается, но не доказано, вертикальная передача кишечной реовирусной инфекции.
Клинические признаки и патология
Пораженные птицы страдают диареей, депрессией, взъерошенными перьями, а также сниженным набором веса [93] и смертностью. Инфекция АРВ может также предрасполагать птиц к вторичным осложнениям или действовать синергетически вместе с другими инфекциями. Сообщалось, что АРВ-инфекция может усиливать кокцидиальную инфекцию у цыплят-бройлеров [85].Также существует возможность синергического действия TRV с другими патогенами у индеек.
В большинстве случаев содержимое кишечника пенистое и водянистое. В некоторых случаях наблюдается атрофия бурсы Фабрициуса [22, 93]. Поражения кишечника состоят из легкой гиперплазии крипт из-за инфильтрации лимфоцитов, гетерофилов и эозинофилов в собственную пластинку и подслизистую оболочку через 11–14 дней после заражения. В сумке Фабрициуса наблюдается умеренное или тяжелое истощение фолликулярных лимфоцитов [22, 93].Помимо атрофии бурсальной сумки, может наблюдаться лимфоидное истощение от легкой до умеренной степени в селезенке и лимфоцитарная инфильтрация в печени, поджелудочной железе, сердце и преджелудках.
Диагноз
Диарея в стаде является признаком кишечной инфекции. Трудно точно определить вирус / возбудитель только на основе диареи, и для подтверждения необходимы лабораторные тесты. Электронная микроскопия может выявить присутствие вирусных частиц размером ~ 70–80 нм в кишечнике инфицированных птиц.Вирус хорошо растет в курином яйце с эмбрионом через желточный мешок или хориоаллантоисную мембрану, а также был выделен в первичных клетках печени куриного эмбриона (CEL), клетках почек куриного эмбриона (CEK) и клетках фибросаркомы японского перепела (QT-35). Из-за фузогенной природы TRV в инокулированных культурах клеток наблюдается характерный цитопатический эффект (ЦПЭ). Для обнаружения антител против реовируса цыплят коммерчески доступны наборы ELISA, но в настоящее время таких специфических наборов для реовируса индейки нет.Недавно мы разработали тест ELISA для обнаружения антител против реовируса индейки. В этом тесте мы клонировали и экспрессировали белок сигма C S1 и использовали его в качестве антигена для обнаружения антител против TRV (неопубликованные данные Mor et al.).
Доступно несколько тестов ОТ-ПЦР, нацеленных на различные генные сегменты реовируса [4, 20, 44, 45, 52, 88]. ОТ-ПЦР в реальном времени — это наиболее чувствительный и специфический тест, с помощью которого можно количественно определить вирусную нагрузку в клинических образцах. Был разработан надежный, сверхчувствительный и точный количественный анализ на АРВ с помощью Light Cycler SYBR Green на основе ОТ-ПЦР в реальном времени, который может обнаруживать 39 копий / мкл геномной РНК АРВ [53].Недавно в нашей лаборатории была разработана высокочувствительная и специфическая ОТ-ПЦР в реальном времени для обнаружения TRV. Мы разработали праймеры и зонды из сегмента генома S4. Тест специфичен для TRV (энтерических вирусов индейки и реовирусов артрита) и может обнаруживать до 10 копий транскрибированной in vitro стандартной вирусной РНК на реакцию (Mor et al. Неопубликованные данные).
Профилактика и контроль
Эффективного лечения реовирусных инфекций не существует. Вирус является повсеместным по своей природе, и из-за его высокой стабильности в окружающей среде трудно избавить птиц и фермы от реовируса.В настоящее время вакцины против реовируса индейки нет, и поэтому биозащита остается единственным способом борьбы с инфекцией.
Ротавирус
Ротавирусы являются важными патогенами, которые вызывают неонатальную диарею у людей, а также у многих животных и птиц. Этот вирус также был обнаружен у коммерческих цыплят-бройлеров, страдающих синдромом короткого роста и задержки роста [72].
Геном
Ротавирусы — это РНК-вирусы, принадлежащие к роду Rotavirus и семейству Reoviridae .Геном ротавируса состоит из 11 сегментов двухцепочечной РНК; РНК окружена тройным слоем икосаэдрического белкового капсида. Каждый сегмент кодирует как минимум один белок. Внешний белковый слой состоит из двух основных нейтрализующих антигенов — вирусного белка 4 (VP4) и вирусного белка 7 (VP7). Вирусный белок 6 (VP6) второго слоя капсида называется групповым антигеном, который используется для обнаружения и классификации ротавирусов на отдельные группы. Кроме того, по крайней мере пять неструктурных белков кодируются геномом ротавируса.
Основываясь на антигенных свойствах VP6, ICTV разделил ротавирусы на пять серологических видов (A – E) и два дополнительных вида (F и G) [81], которые также называются группами ротавирусов. Известно, что группы A, B и C инфицируют людей и животных, а группы D, E, F и G заражают только животных, в основном птиц [50, 63, 102]. Недавно было сообщено о потенциально новом виде RV, ротавирусе H [ 64 ]. Эти авторы утверждают, что значение отсечения аминокислотной последовательности 53% позволяет дифференцировать различные виды ротавирусов.
В отличие от ротавирусов птиц группы A, ротавирусы группы D не могут размножаться в клетках MA104 [ 25 ] и в основном идентифицируются по схеме электрофоретической миграции 5: 2: 2: 2 их сегментов генома, что значительно отличается от паттерна 5: 1: 3: 2 ротавирусов птичьей группы А. Однако классификация штамма ротавируса только на основе его профиля миграции РНК является проблематичной из-за возможного возникновения перестройки генома. В отличие от ротавирусов группы А, которые преимущественно инфицируют клетки двенадцатиперстной кишки, вирусы группы D имеют склонность к тощей и подвздошной кишке.
Ротавирус индейского происхождения отличается от ротавируса куриного происхождения [58]. Например, сообщалось, что гомология аминокислотной последовательности гена VP7 штамма Ty-1 (индейка) и гена Ch-2 VP7 птиц составляет только 70% [58]. Вариабельные эпитопные области A, B и C Ty-1 были уникальными по сравнению с таковыми у Ch-2 и других штаммов, представляющих серотипы 14G. Низкая гомология (53%) областей A и C Ty-1 и Ch-2 предполагает, что штамм Ty-1 может иметь другой серотип, чем эталонный штамм G7 Ch-2.Сравнение последовательностей различных генов (VP4, VP6, VP7, NSP5) ротавирусов индейки и кур показало, что среди ротавирусов птичьей группы А может происходить межвидовая передача и реассортация [87].
Заболеваемость и распространение
Ротавирусы распространены по всему миру и были зарегистрированы как у здоровых, так и у пораженных энтеритом индеек [44, 45, 69, 71, 75, 77, 99]. Периодический мониторинг товарных стад индейки выявил циркуляцию четырех групп (на основе анализа последовательности гена NSP4) ротавирусов.Среди четырех групп ротавирусов было 96,1–97,5 и 97,5–99,55% сходства на уровне нуклеотидов и аминокислот соответственно [77]. Ротавирусы присутствовали до размещения, а также после размещения до 12-недельного возраста [77]. В другом исследовании, в котором участвовали 33 стада индейки и 43 стада кур, ротавирусы были обнаружены в 46% стадах цыплят и 70% стадах индейки [75]. Мы также сообщали о присутствии ротавирусов в практически здоровых стадах производителей индейки с максимальной встречаемостью до 5-недельного возраста [45].В другом исследовании 93% стад, пораженных PES, были положительными на ротавирусы [44]. В контрольном исследовании Mor et al. [68] сообщили, что шесть (7,5%) пулов образцов фекалий были положительными на ротавирус в стадах, пораженных LTS, в то время как 13 (33%) пулов образцов были положительными из стада без LTS.
Выявление ротавирусов выше у молодых цыплят, и уровень обнаружения снижается с возрастом. При наблюдении PES и LTS максимальное количество цыплят было положительным на ротавирус в возрасте 2–5 недель с последующим уменьшением количества положительных образцов в возрасте 8–9 недель [44, 45, 68].Было показано, что праймеры на основе высококонсервативной области генома ротавируса (в частности, неструктурных белков NSP3 или NSP4) подходят для идентификации ротавируса в различных клинических образцах, образцах животных или окружающей среды с помощью обычных или в реальном времени RT- ПЦР [51, 84]. В исследовании, проведенном в Европе, ротавирусы кур и индеек были идентифицированы с использованием двух разных методов: электрофореза в полиакриламидном геле с ОТ-ПЦР и анализов ОТ-ПЦР в реальном времени, специфичных для ротавирусов групп A и D [71].Эти результаты показывают, что необходимы дальнейшие исследования для точного определения типов ротавирусов, циркулирующих в стадах индейки в различных географических точках.
Передача
Фекальный оральный путь является основным путем горизонтальной передачи. Вирус может выжить в окружающей среде в течение длительного времени, и, следовательно, механическая передача может играть роль в его передаче. Есть подозрение на вертикальную передачу ротавируса, но это пока не подтверждено. Например, Pantin-Jackwood et al.[77] обнаружили ротавирусы в 50% проб, исследованных до помещения птицы. Основываясь на этой информации, авторы предположили, что ротавирус может передаваться вертикально. Мы также обнаружили ротавирусы у 2-дневных коммерческих и родительских домашних птиц [45, 47], что указывает на возможность вертикальной передачи ротавируса.
Также описана межвидовая передача ротавируса [87, 103]. В одном исследовании ротавирусы крупного рогатого скота группы А (n = 6) были обнаружены в кишечном содержимом индюков [2].Электроферотипы показали картину миграции, идентичную вирусу диареи телят в Небраске, а полная филогения гена NSP4 показала, что все шесть штаммов сегрегированы по генотипу E2, что подтверждает их принадлежность к ротавирусу крупного рогатого скота. Ротавирусы птиц не обнаружены у людей, и ротавирусы человека не содержат последовательностей ротавирусов птиц [87], что указывает на низкую вероятность заражения человека ротавирусами птиц.
Клинические признаки и патология
Тупость, депрессия, водянистый помет, потеря аппетита, склеивание вентиляционных отверстий и снижение прибавки в весе — постоянные признаки у пораженных птиц.Смертность может присутствовать или отсутствовать, а заболеваемость может варьироваться. Ротавирусная инфекция кур-несушек может привести к снижению яйценоскости.
В большинстве случаев наблюдается пенистое и водянистое кишечное содержимое. Может наблюдаться бледность кишечного тракта и вздутие слепой кишки с пенистым или непенистым жидким содержимым. Другими крупными патологическими проявлениями являются обезвоживание, задержка роста и пастообразные выделения. Также отмечаются мультифокальные и поверхностные эрозии двенадцатиперстной и тощей кишки. Наиболее частыми микроскопическими изменениями являются гиперцеллюлярность собственной пластинки из-за инфильтрации гетерофилов, эозинофилов и мононуклеарных клеток, а также атрофия ворсинок.Сканирующая электронная микроскопия показывает шероховатость поверхности ворсинок, нарушение нормальной морфологии ворсинок и потерю микроворсинок в клетках, расположенных на кончиках ворсинок. У цыплят разные штаммы ротавирусов могут иметь разные предпочтительные сайты репликации. Например, McNulty et al. [66] сообщили, что у экспериментально инфицированных кур ротавирус группы A имел сродство к двенадцатиперстной кишке, тогда как ротавирус группы D предпочитал тощую кишку и подвздошную кишку.
Диагноз
Диарея, обезвоживание и снижение веса стада свидетельствуют о кишечной инфекции.ЭМ кишечного содержимого и фекалий выявляет присутствие вирусных частиц размером ~ 70 нм (приблизительно 50–70 нм) с характерным «колесом» внешнего вида. Несмотря на то, что вирус трудно изолировать, он все же растет в клетках MA104 при обработке трипсином. Тест коагглютинации использовался для обнаружения ротавирусов у индеек и оказался простым и быстрым скрининговым тестом. Электрофорез в полиакриламидном геле обычно используется для определения характерной картины миграции 11 сегментов генома ротавируса и почти так же чувствителен, как ЕМ.
Для обнаружения вируса доступно несколько ОТ-ПЦР и ОТ-ПЦР в реальном времени, нацеленных на различные сегменты генома ротавируса [8, 21, 77]. Недавно было сообщено о высокочувствительной и специфической одностадийной ОТ-ПЦР в реальном времени, нацеленной на неструктурный белок 4 (NSP4), с внутренней системой контроля для обнаружения ротавирусов индейки [1].
Профилактика и борьба
Специальных мер борьбы с ротавирусными инфекциями не существует. Получение цыплят из источников, свободных от ротавирусов, и принятие надлежащих мер биобезопасности — это способы минимизировать заражение ротавирусом в стаде.
Астровирус
Астровирус индейки (TAstV) был впервые обнаружен у индюшат в возрасте 6–11 дней с диареей и повышенной смертностью в Великобритании. В США TAstV был обнаружен у индюшат и позже был назван TAstV-1. Впоследствии TAstV, связанный с PEMS, был выделен и охарактеризован. Этот вирус генетически и иммунологически отличался от ранее описанного изолята США, TAstV-1, и был назван TAstV-2 [56, 86]. Помимо TAstV-1 и TAstV-2, астровирус куриного происхождения i.е. вирус птичьего нефрита (ВНВ) обнаружен также у индеек [26, 75]. ANV сначала был описан как небольшой круглый вирус, затем как пикорнавирус, а позже был определен как астровирус [40]. Таким образом, у индеек обнаружено три типа астровирусов (TAstV-1, TAstV-2 и ANV). Однако данные о последовательностях предполагают, что в природе может присутствовать гораздо больше типов [94]. В 1980-х годах все обнаруженные астровирусы были строго ограничены желудочно-кишечным трактом домашних птиц, и не было никаких признаков системного заболевания.Однако TAstV также был обнаружен в тимусе и сумке, что указывает на возможное влияние на иммунную систему [86]. Сообщалось также об обнаружении астровирусов у кур и индюшат, страдающих кишечными и / или опорно-двигательными расстройствами (теносиновит) [26, 82].
Геном
Астровирусы — это РНК-вирусы без оболочки диаметром 25–35 нм, принадлежащие к роду Avastrovirus семейства Astroviridae . Геном астровируса состоит из 6,8–7 РНК.9 kb и имеет 5′-UTR, за которым следуют три ORF, 3′-UTR и поли-A-хвост. ORF-1a кодирует сериновую протеазу и другие неструктурные белки, ORF-1b для РНК-зависимой РНК-полимеразы и ORF-2 для капсидного белка. Общая длина геномов трех астровирусов, а именно. TAstV-1 (7003 нт), TAstV-2 (7325 нт) и ANV (6927 нт) варьируются; эта длина не включает хвост поли-А [55].
Большинство исследований по обнаружению и характеристике TAstV было проведено молекулярными методами с использованием праймеров, специфичных для гена капсида или полимеразы.Ряд исследований показал вариации на уровне нуклеотидов и аминокислот в гене капсида и / или гене полимеразы TAstVs [13, 44–46, 78, 79, 97]. Сообщается о высоком уровне генетической изменчивости среди астровирусов индейки из стад индейки в США [78]. Идентичность нуклеотидной последовательности в гене капсида, связанная с серотипом и вирусным патогенезом, была всего лишь 69%, но ген полимеразы был более консервативным с 86–99% идентичностью нуклеотидов. Недавно филогенетический анализ белка-предшественника капсида выявил обширные вариации в аминокислотных последовательностях TAstV-2 (81.5–100%), но не в TAstV-1 (96,2–100%) [79]. Мы также обнаружили вариации на уровне аминокислот в гене капсида TAstV-2 [44, 46]. Эти изменения предполагают, что в природе могут существовать различные серотипы астровируса индейки. Для лучшего понимания серотипов TAstV, циркулирующих у индеек, необходимы исследования полных последовательностей генов капсида из явно здоровых и пораженных PEC стад из разных географических регионов. Изменения гена капсида из-за мутаций или рекомбинации могут влиять на антигенность и патогенность вирусов и, следовательно, иметь практическое значение для методов обнаружения вирусов, эпидемиологических исследований и разработки потенциальных вакцин против астровирусных инфекций.
Заболеваемость и распространение
Был проведен ряд исследований для выявления присутствия TAstV в стадах индейки с использованием различных методов. Ранее вирусы в образцах кишечника выявлялись с помощью прямой или иммунной ЭМ. Однако в большинстве недавних исследований используются молекулярные инструменты, такие как ОТ-ПЦР и секвенирование, для обнаружения различных астровирусов [36, 56, 57, 108]. В ходе исследования восьми промышленных предприятий по выращиванию индейки и одного исследовательского подразделения в США, 89,5% из 96 образцов содержимого кишечника были положительными на астровирус с помощью ОТ-ПЦР [77].В исследовании, проведенном в Миннесоте, 84% из 43 стай индеек, пораженных PES, оказались положительными на TAstV-2 с помощью ОТ-ПЦР [44]. Эти образцы не тестировались на наличие TAstV-1 и ANV, поэтому нельзя исключать присутствие TAstV-1 и ANV в этих образцах. Ряд других исследований также выявил высокую распространенность TAstV в стадах индейки в США [75, 77, 78].
Исследования распространенности TAstV также проводились в других странах, где выращивают индюшат [17, 27, 62, 70, 74]. Да Силва и др.[17] сообщили о TAsV у молодых цыплят, пораженных PEC в Бразилии. В другом исследовании, проведенном в Бразилии, TAsV-1, TAstV-2 и ANV были идентифицированы в 64,5, 44,7 и 35,5% образцов кишечника соответственно [70]. Из них TAstV-1 и TAstV-2 были более распространены в стадах индеек, пораженных энтеритом. Domanska-Blicharz et al. [27] сообщили, что TAstV-2 является наиболее распространенным вирусом в Польше; 30 из 77 стад индейки (38,9%) были положительными. TAstV-1 был обнаружен только в девяти стадах, тогда как ANV — в одном стаде. Эти три вируса были обнаружены либо как одиночные, либо как смешанные инфекции.Тестирование 23 образцов кишечника от откормленных индюков с клиническими признаками энтерита выявило присутствие TAstV-2 в 17 образцах в Хорватии [62].
TAstV были обнаружены не только в стадах индюшат, пораженных энтеритом, но также и в практически здоровых стадах индюшат [45, 68, 78]. Jindal et al. [45] протестировали 193 объединенных образца фекалий практически здоровых индюшат-производителей с помощью ОТ-ПЦР и обнаружили, что 47,2% оказались положительными на TAstV-2.
Передача
Передача астровирусов фекально-оральным путем.Здоровые птицы могут подвергаться воздействию астровируса либо при прямом контакте с инфицированными фекалиями, либо с материалами, загрязненными фекалиями. Считается, что астровирусные инфекции видоспецифичны, но недавние сообщения указывают на межвидовую передачу [23, 101]. Так, TAstV-2 и ANV были обнаружены у уток ( Anas platyrhynchos domesticus ) [9]. Межвидовая передача и адаптация вируса к новым хозяевам или совместное инфицирование одного и того же хозяина разными астровирусами может привести к появлению новых астровирусов, которые инфицируют животных или обладают зоонозным потенциалом [24].
Клинические данные и патология
Как и при рота- и реовирусных инфекциях, астровирусы также вызывают диарею, обезвоживание и задержку роста. Сообщалось о подобных наблюдениях диареи и задержки роста у экспериментальных цыплят, инокулированных материалом, содержащим TAstV [48, 49, 76]. Были предприняты экспериментальные исследования, чтобы установить тип клинического заболевания, вызываемого TAstV, имеющими вариантные капсиды, или те, которые различались по источнику происхождения. Например, Pantin-Jackwood et al.[76] изучали патогенность трех различных вирусов TAstV-2 у индюков, свободных от специфических патогенов, и обнаружили, что вирусы TAstV-2 с вариантными капсидами вызывают подобное кишечное заболевание у молодых индеек. Mor et al. [67] обнаружили, что TAstV-2 от птиц PES был более патогенным, чем TAstV-2 от практически здоровых домашних птиц. Астровирус может потенциально нарушать иммунный ответ, тем самым повышая восприимчивость индеек к вторичным бактериальным инфекциям [80].
Обезвоживание, вздутие кишечника, наполненное водянистым содержимым и непереваренной пищей, и расширенная слепая кишка с пенистым содержимым — это результаты вскрытия.Может возникнуть атрофия бурсальной сумки от легкой до умеренной, а также снижение тонуса кишечника. Гистологическими находками являются умеренный некроз эпителия, инфильтраты собственной пластинки и легкая гиперплазия крипт.
Диагноз
Диарея, обезвоживание и снижение веса являются признаками кишечной инфекции. Электронная микроскопия с отрицательным контрастом выявляет вирусные частицы с пятью или шестью выступами на поверхности, напоминающими звезду. Звездообразная морфология не всегда сохраняется или четко различима.По этой причине вирусы AstV от EM можно спутать с пикорнавирусами, пикорнавирусами, энтеровирусами и энтероподобными вирусами. Во-вторых, домашние птицы могут демонстрировать клинические признаки и патологию на более поздней стадии астровирусной инфекции, но в кишечном содержимом может не быть обнаруживаемых астровирусных частиц. В такой ситуации, даже если заболевание возникло, результаты анализов могут быть отрицательными. Молекулярные методы были использованы для обнаружения и дальнейшей характеристики астровирусов. В большинстве анализов на основе ОТ-ПЦР используются праймеры, специфичные для полимеразы или гена капсида TAstV [56, 57, 96].
Смешанные вирусные инфекции
Хотя отдельные вирусы были обнаружены в стадах индейки, пораженных энтеритом, большинство исследований указывают на присутствие двух или более энтеральных вирусов [44, 45, 62, 68, 70, 75, 77, 105, 107 ]. Периодический мониторинг восьми промышленных стад индейки в США выявил постоянное присутствие TAStV и ротавируса [77]. Из 96 протестированных образцов 89,5% были положительными на астровирус и 67,7% на ротавирус. Все стада были отрицательными на TCoV, реовирус и аденовирус группы 1 во всех точках отбора проб.В другом исследовании присутствие астровирусов было зарегистрировано в 100% протестированных стад индейки [75]. TAstV-2 и TAstV-1 были обнаружены в 100 и 15,4% стад индейки соответственно. Кроме того, 12,5% стад индейки были положительными на ANV. Ротавирусы и реовирусы присутствовали в 69,7 и 45,5% стад соответственно, при этом коронавирус и аденовирус не обнаружены.
Кишечное содержимое 43 стад PES в Миннесоте было проверено на наличие энтеральных вирусов методом RT-PCR.Все 43 стада оказались положительными на ротавирус, TAstV-2 или реовирус. Во всех случаях вирусы были обнаружены либо по отдельности, либо в комбинации из 2 или 3. Восемь стай (19%) были положительными на один вирус, а остальные 35 (81%) имели комбинацию вирусов. Пятнадцать стад оказались положительными на все три вируса. Ни одно из протестированных стад не было признано положительным на TCoV [44]. Исследования, проведенные в Бразилии, показали наличие 69,7% множественных вирусов в образцах кишечника индеек. Образцы кишечника чаще всего содержали 3–4 вируса на образец, причем вирусы из семейства астровирусов (TAstV-1, TAstV-2 или ANV, или их комбинация) присутствовали во всех образцах, которые дали положительный результат на вирусы [70 ].
Возможно, что комбинация вирусов может вызвать более серьезные побочные эффекты у домашних птиц, чем отдельные вирусы. В одном исследовании индюшатина, инокулированная комбинацией TAstV-2, реовируса индейки и ротавируса, имела значительно меньшую массу тела, чем контрольная группа [91]. В экспериментальных исследованиях мы также сообщали, что пероральная инокуляция цыплят материалом PES (положительным по ротавирусу, TAstV-2 и Salmonella) приводила к значительной задержке роста на срок до 50 дней после инокуляции, и ни в какой момент после инокуляции не наблюдалось. веса оспариваемых птиц сходятся с весом контрольных.Содержимое кишечника было водянистым и пенистым / пенистым (рис.,) У цыплят, инокулированных материалом PES [49]. Точно так же индюшатины, инокулированные содержимым кишечника птиц, пораженных LTS, имели значительно меньшую массу тела в возрасте 20 недель по сравнению с индюшатинами, инокулированными содержимым кишечника птиц без LTS [68]. Такая степень замедления роста из-за энтерита домашней птицы может привести к значительным экономическим потерям для производителей индейки, поскольку пораженная птица может не достичь ожидаемого веса при продаже.На уровне фермы нельзя исключать роль агентов, отличных от этих четырех распространенных вирусов (TAstV, ротавирус, реовирус и коронавирус), в возникновении и / или повышении серьезности кишечных вирусов. Кроме того, факторы питания и менеджмент также могут играть роль в возникновении энтерита у стада. Такие шаги, как улучшение методов лечения и противомикробная терапия для минимизации потерь, имели ограниченный успех [5]. Вакцины против кишечных вирусов в настоящее время нет.
Водянистое и пенистое содержимое слепой кишки птицы, инокулированной перорально кишечным материалом птиц, пораженных синдромом энтерита домашних птиц
Водянистое и пенистое содержимое слепой кишки птицы, инокулированной перорально кишечным материалом птиц, пораженных синдромом энтерита домашних птиц.Слепая кишка разрезана
Кишечные вирусы обычно обнаруживаются с помощью ЭМ, серологии и ПЦР. Большинство из этих вирусов трудно вырастить в зародышевых яйцах курицы / индейки и клеточных культурах [95]. Таким образом, было разработано несколько анализов одиночной и множественной RT-PCR (mRT-PCR) для обнаружения астровируса индейки, ротавируса индейки, коронавируса индейки и реовируса индейки [21, 43, 56, 77, 89, 92]. Мультиплексные анализы RT-PCR экономят время и деньги, поскольку два или более вируса могут быть обнаружены в одной реакции.
Общие выводы и направления на будущее
Кишечные вирусы (TAstV, ротавирус, реовирус и коронавирус) широко распространены в стадах индейки, но их клиническое значение неясно. Полное влияние этих вирусов на производительность стада требует тщательного изучения. Хотя кишечные заболевания, вызванные вирусами, являются экономическим бременем для фермеров, выращивающих индейки, ни один вирус не появился в качестве вероятного возбудителя, чтобы можно было сформулировать соответствующие меры по профилактике и контролю [18].В нескольких исследованиях одновременно были обнаружены два или более разных вируса, что указывает на то, что несколько вирусов могут участвовать в патогенезе энтерита. Преобладание вирусов одного и того же рода в кишечнике как здоровых, так и плохо функционирующих птиц указывает на существование разных патотипов вируса. Необходимы дальнейшие исследования, чтобы определить обоснованность этих утверждений.
С появлением молекулярных диагностических тестов и секвенирования наше понимание кишечных вирусов расширилось.Также существует вероятность того, что наши текущие методы не могут обнаружить некоторые еще неизвестные вирусы / патогены, которые могут иметь последствия для энтерита индейки. Доступность и использование высокопроизводительных методов пиросеквенирования нуклеиновых кислот нового поколения, таких как секвенирование Illumina, может выявить последовательности агентов, о которых ранее не сообщалось у индеек. Метагеномные исследования РНК-вирусов в кишечнике индеек, страдающих кишечным заболеванием, выявили присутствие последовательностей вирусов дцРНК ( Reoviridae и пикобирнавирусы) и вирусов оцРНК ( Caliciviridae , Leviviridae , Picornaviridae Astroviridae и Picornaviridae ). [18].Обнаружение неизвестных вирусов методом метагеномики проложит путь к разработке методов диагностики этих вирусов. Кроме того, необходимо определить, почему одни и те же типы вируса обнаруживаются в стадах индейки, пораженных энтеритом, а также в явно здоровых стадах. Полное понимание роли кишечных вирусов и других патогенов в кишечных болезнях индеек будет иметь большое значение при разработке профилактических и контрольных мер.
Ссылки
1. Акимкин В., Биндель Ф., Хоферер М., Стинг Р., Полли Б., Ханель А., Хафез Х.М.Одношаговая RT-qPCR с системой внутреннего контроля для обнаружения ротавирусов индейки в образцах фекалий. J Virol Methods. 2011. 177 (1): 112–117. [PubMed] [Google Scholar] 2. Асано К. М., Грегори Ф., Соуза С. П., Ротава Д., Оливейра Р. Н., Вильярреал Л. Я., Рихтценхайн Л. Дж., Брандао ЧП. Ротавирус крупного рогатого скота у индеек с энтеритом. Птичий Дис. 2011; 55 (4): 697–700. [PubMed] [Google Scholar] 3. Трепет О.О., Али А., Элайш М., Ибрагим М., Мурджа М., Пантин-Джеквуд М., Саиф Ю.М., Ли К.В. Влияние коронавирусной инфекции на репродуктивную способность кур индюков.Птичий Дис. 2013; 57: 650–656. [PubMed] [Google Scholar] 4. Баньяи К., Дандар Э., Дорси К.М., Мато Т., Паля В. Геномное созвездие нового штамма ортореовируса птиц, связанного с синдромом низкорослости у бройлеров. Гены вирусов. 2011; 42: 82–89. [PubMed] [Google Scholar]5. Барнс Х. Дж., Гай Дж. С.. Синдром энтерита-смертности птицы. В: Саиф Ю.М., Барнс Х.Дж., Глиссон-младший, Фэдли А.М., Макдугалд Л.Р., Суэйн Д.Е., редакторы. Болезни птицы. 11-е изд. Эймс: Издательство государственного университета Айовы; 2003. С. 1171–1180.
6. Barnes HJ, Guy JS, Vaillancourt JP. Комплекс энтерита птицы. Rev Sci Tech. 2000. 19: 565–588. [PubMed] [Google Scholar] 7. Бенавенте Дж., Мартинес-Костас Дж. Птичий реовирус: структура и биология. Virus Res. 2007. 123: 105–119. [PubMed] [Google Scholar] 8. Безерра Д.А., да Силва Р.Р., Кайано Дж. Х., Сильвестр Р. В., де Соуза Оливейра Д., Линьярес С., Габбай Ю. Б., Маскареньяс Дж. Д.. Обнаружение ротавируса группы D птиц с использованием полимеразной цепной реакции для гена VP6. J Virol Methods. 2012; 185: 189–192. [PubMed] [Google Scholar] 9.Bidin M, Bidin Z, Majnaric D, Tisljar M, Lojkic I. Циркуляция и филогенетическая взаимосвязь астровирусов куриного и индейского происхождения, обнаруженных у домашних уток ( Anas platyrhynchos domesticus ) Avian Pathol. 2012. 41 (6): 555–562. [PubMed] [Google Scholar] 10. Боделон Дж., Лабрада Л., Мартинез-Костас Дж., Бенавенте Дж. Сегмент генома реовируса птиц S1 представляет собой функционально трицистронный ген, который экспрессирует один структурный и два неструктурных белка в инфицированных клетках. Вирусология. 2001; 290: 181–191.[PubMed] [Google Scholar] 11. Бреслин Дж. Дж., Смит Л. Г., Барнс Г. Дж., Гай Дж. С.. Сравнение процедур выделения вируса, иммуногистохимии и полимеразной цепной реакции с обратной транскриптазой для обнаружения коронавируса индейки. Птичий Дис. 2000. 44: 624–631. [PubMed] [Google Scholar] 12. Бангер А. Н., Чакон Дж. Л., Джонс Р. К., Феррейра А. Дж.. Обнаружение и молекулярная характеристика гена 3 и 5 коронавируса индейки у индюшат с тяжелым энтеритом в Бразилии. Птичий Дис. 2009. 53: 356–362. [PubMed] [Google Scholar] 13.Канелли Э., Кордиоли П., Барбьери И., Кателла А., Пеннелли Д., Церути Р., Морено А., Лавацца А. Астровирусы как возбудители энтерита домашней птицы: генетическая характеристика и лонгитюдные исследования в полевых условиях. Птичий Дис. 2012; 56: 173–182. [PubMed] [Google Scholar] 14. Cardoso TC, Castanheira TL, Teixeira MC, Rosa AC, Hirata KY, Astolphi RD, Luvizotto MC. Валидация иммуногистохимического анализа для обнаружения коронавируса индейки: быстрый и простой инструмент для скрининга в условиях ограниченных ресурсов. Poult Sci.2008. 87 (7): 1347–1352. [Бесплатная статья PMC] [PubMed] [Google Scholar] 15. Чен Ю.Н., Ву СС, Брайан Т., Хупер Т., Шредер Д., Лин Т.Л. Специфическая полимеразная цепная реакция обратной транскрипции в режиме реального времени для обнаружения и количественного определения РНК коронавируса индейки в тканях и фекалиях индеек, инфицированных коронавирусом индейки. J Virol Methods. 2010. 163: 452–458. [Бесплатная статья PMC] [PubMed] [Google Scholar] 16. Калвер Ф.А., Бриттон П., Кавана Д. Обнаружение с помощью ОТ-ПЦР птичьих коронавирусов галлиформных птиц (курица, индейка, фазан) и у попугая.Методы Мол биол. 2008; 454: 35–42. [Бесплатная статья PMC] [PubMed] [Google Scholar] 17. Да Силва SEL, Бонетти AM, Petrocelli ATM, Ferrari HF, Luvizotto MCR, Cardoso TC. Обнаружение астровируса индейки у молодняка, пораженного комплексом энтерита домашней птицы, в Бразилии. J Vet Med Sci. 2008. 70 (6): 629–631. [PubMed] [Google Scholar]19. Дэй Дж. М., Гондер Э., Дженнингс С., Ривес Д., Тилли Б., Вуминг Б. Молекулярная характеристика кишечных коронавирусов индейки, циркулирующих в США в 2012 году. Документ, представленный на ежегодном собрании Американской ассоциации патологоанатома в Чикаго, штат Иллинойс.2013.
20. Day JM, Pantin-Jackwood MJ, Spackman E. Последовательный и филогенетический анализ сегмента S1 генома реовирусов индейского происхождения. Гены вирусов. 2007. 35 (2): 235–242. [PubMed] [Google Scholar] 21. Дэй JM, Spackman E, Пантин-Джеквуд MJ. Мультиплексный ОТ-ПЦР-тест для дифференциальной идентификации астровируса индейки типа 1, астровируса индейки типа 2, астровируса цыпленка, вируса птичьего нефрита и птичьего ротавируса. Птичий Дис. 2007. 51: 681–684. [PubMed] [Google Scholar] 22. Дэй JM, Spackman E, Пантин-Джеквуд MJ.Иммунная дисфункция, вызванная реовирусом индейского происхождения, у свободных от конкретных патогенов и промышленных индюшат. Птичий Дис. 2008. 52 (3): 387–391. [PubMed] [Google Scholar] 23. Де Баттисти С., Сальвиато А., Йонассен С.М., Тоффан А., Капуа I, Каттоли Г. Генетическая характеристика астровирусов, обнаруженных у цесарок ( Numida meleagris ), выявляет отдельный генотип и предполагает межвидовую передачу между индейкой и цесаркой. Arch Virol. 2012. 157 (7): 1329–1337. [PubMed] [Google Scholar] 24. Де Бенедиктис П., Шульц-Черри С., Бернем А., Каттоли Г.Астровирусные инфекции у людей и животных — молекулярная биология, генетическое разнообразие и межвидовая передача. Заразить Genet Evol. 2011. 11 (7): 1529–1544. [Бесплатная статья PMC] [PubMed] [Google Scholar] 25. Девитт CM, Рейнольдс DL. Характеристика ротавирусов группы D. Птичий Дис. 1993; 37: 749–755. [PubMed] [Google Scholar] 26. de Wit JJ, ten Dam GB, van de Laar JMAM, Biermann Y, Verstegen I, Edens F, Schrier CC. Обнаружение и характеристика нового астровируса у кур и индеек с кишечными расстройствами и нарушениями опорно-двигательного аппарата.Avian Pathol. 2011; 40: 453–461. [PubMed] [Google Scholar] 27. Доманска-Бличарц К., Серока А., Минта З. Годовое молекулярное исследование астровирусной инфекции у индеек в Польше. Arch Virol. 2011. 156 (6): 1065–1072. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Франка М., Креспо Р., Чин Р., Вулкок П., Шивапрасад Х.Л. Ретроспективное исследование миокардита, ассоциированного с реовирусом, у индеек. Птичий Дис. 2010. 54 (3): 1026–1031. [PubMed] [Google Scholar] 30. Gomaa MH, Yoo D, Ojkic D, Barta JR. Распространенность коронавируса индейки у североамериканских индеек определяется с помощью недавно разработанного иммуноферментного анализа на основе рекомбинантного антигена.Clin Vaccine Immunol. 2008. 15 (12): 1839–1844. [Бесплатная статья PMC] [PubMed] [Google Scholar] 31. Gomaa MH, Yoo D, Ojkic D, Barta JR. Использование рекомбинантного полипептида-шипа S1 для разработки ELISA-анализа антител, специфичных к TCoV. Vet Microbiol. 2009. 138 (3–4): 281–288. [Бесплатная статья PMC] [PubMed] [Google Scholar] 32. Gomaa MH, Yoo D, Ojkic D, Barta JR. Заражение патогенным изолятом коронавируса индейки отрицательно влияет на показатели роста и морфологию кишечника молодняка индейки в Канаде. Avian Pathol.2009. 38: 279–286. [PubMed] [Google Scholar] 33. Гомес Д.Е., Хирата К.Ю., Сахеки К., Роза А.С., Лувизотто М.С., Кардосо ТК. Патология и тканевое распределение коронавируса индейки у экспериментально инфицированных цыплят и индюшат. J Comp Pathol. 2010. 143 (1): 8–13. [Бесплатная статья PMC] [PubMed] [Google Scholar] 34. Гай JS. Коронавирусный энтерит в Турции. В: Саиф Ю.М., Барнс Х.Дж., Глиссон-младший, Фэдли А.М., Макдугалд Л.Р., Суэйн Д.Е., редакторы. Болезнь птицы. 11. Эймс: Издательство государственного университета Айовы; 2003. С. 300–307. [Google Scholar] 35.Гай JS. Коронавирусный энтерит в Турции. В: Саиф YM, Fadly AM, Glisson JR, McDougald LR, Nolan LK, Swayne DE, редакторы. Болезни птицы. 12. Эймс: специалист по издательскому делу Wiley-Blackwell; 2008. С. 330–338. [Google Scholar] 36. Guy JS, Miles AM, Smith L, Fuller FJ, Schultz-Cherry S. Антигенная и геномная характеристика энтеровирусоподобного вируса индейки (Северная Каролина, изолят 1988 г.): идентификация вируса как астровируса индейки 2. Avian Dis. 2004. 48 (1): 206–211. [PubMed] [Google Scholar] 37.Гай Дж. С., Смит Л. Г., Бреслин Дж. Дж., Пакпиньо С. Разработка конкурентного иммуноферментного анализа для обнаружения антител к коронавирусу индейки. Птичий Дис. 2002. 46 (2): 334–341. [PubMed] [Google Scholar] 38. Гай Дж. С., Смит Л. Г., Бреслин Дж. Дж., Вайланкур Дж. П., Барнс Х. Дж. Высокая смертность и подавление роста, экспериментально вызванные у молодых индеек двойным заражением энтеропатогенным вирусом Escherichia coli и коронавирусом индейки. Птичий Дис. 2000. 44: 105–113. [PubMed] [Google Scholar] 39.Heggen-Peay CL, Qureshi MA, Edens FW, Sherry B, Wakenell PS, O’Connell PH, Schat KA. Выделение реовируса от синдрома энтерита и смертности домашних птиц и его патогенность у индюшат. Птичий Дис. 2002. 46 (1): 32–47. [PubMed] [Google Scholar] 40. Имада Т., Ямагути С., Масе М, Цукамото К., Кубо М., Морока А. Вирус птичьего нефрита (ANV) как новый член семейства Astroviridae и конструирование инфекционной кДНК ANV. J Virol. 2000; 74: 8487–8493. [Бесплатная статья PMC] [PubMed] [Google Scholar] 41.Исмаил М.М., Тан А.Ю., Саиф Ю.М. Патогенность коронавируса индейки у индеек и кур. Птичий Дис. 2003. 47: 515–522. [PubMed] [Google Scholar] 42. Джеквуд М.У., Бойнтон Т.О., Хилт Д.А., Маккинли Е.Т., Киссинджер Дж.С., Патерсон А.Х., Робертсон Дж., Лемке С., МакКолл А.В., Уильямс С.М., Джеквуд Дж. В., Берд Л.А. Возникновение коронавируса группы 3 в результате рекомбинации. Вирусология. 2010; 398: 98–108. [Бесплатная статья PMC] [PubMed] [Google Scholar] 43. Джиндал Н., Чандер И., Патнаяк Д.П., Мор СК, Зиглер А.Ф., Гоял С.М. Мультиплексная RT-PCR для обнаружения астровирусов, ротавирусов и реовирусов у индеек.Птичий Дис. 2012; 56: 592–596. [PubMed] [Google Scholar] 44. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М. Выявление и молекулярная характеристика кишечных вирусов синдрома энтерита домашней птицы у индеек. Poult Sci. 2010. 89: 217–226. [Бесплатная статья PMC] [PubMed] [Google Scholar] 45. Джиндал Н., Патнаяк Д.П., Чандер Ю., Зиглер А.Ф., Гоял С.М. Обнаружение и молекулярная характеристика кишечных вирусов у племенных индюков. Avian Pathol. 2010; 39: 53–61. [PubMed] [Google Scholar] 46. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М.Сравнение последовательностей капсидных генов астровируса-2 индейки у индюшат, пораженных синдромом энтерита, и у практически здоровых индюшат. Arch Virol. 2011; 156: 969–977. [PubMed] [Google Scholar] 47. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Ретроспективное исследование синдрома энтерита домашней птицы в Миннесоте. Птичий Дис. 2009; 53: 268–275. [PubMed] [Google Scholar] 48. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Экспериментальное воспроизведение синдрома энтерита домашней птицы: клинические данные, реакция роста и микробиология.Poult Sci. 2009; 88: 949–958. [Бесплатная статья PMC] [PubMed] [Google Scholar] 49. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Продолжительность угнетения роста и выделения патогенов при синдроме энтерита экспериментально воспроизведенных индюшат. Птичий Дис. 2009; 53: 517–522. [PubMed] [Google Scholar] 50. Johne R, Otto P, Roth B, Löhren U, Belnap D, Reetz J, Trojnar E. Анализ последовательности сегмента генома, кодирующего VP6, ротавирусов птичьей группы F и G. Вирусология. 2011; 412: 384–391. [PubMed] [Google Scholar] 51.Джотикумар Н, Канг Г., Хилл ВР. Широкореактивный анализ TaqMan для обнаружения ротавируса в режиме реального времени с помощью ОТ-ПЦР в клинических образцах и образцах окружающей среды. J Virol Methods. 2009. 155: 126–131. [PubMed] [Google Scholar] 52. Капчинский Д.Р., Селлерс Х.С., Симмонс В., Шульц-Черри С. Анализ последовательности гена S3 из реовируса индейки. Гены вирусов. 2002. 25 (1): 95–100. [Бесплатная статья PMC] [PubMed] [Google Scholar] 53. Кэ GM, Cheng HL, Ke LY, Ji WT, Chulu JL, Liao MH, Chang TJ, Liu HJ. Разработка количественного светового циклера RT-PCR в реальном времени для обнаружения птичьего реовируса.J Virol Methods. 2006; 133 (1): 6–13. [PubMed] [Google Scholar] 54. King AMQ, Lefkowiz E, Adams M, Carstens E. Таксономия вирусов: девятый доклад международного комитета по таксономии вирусов. Уолтем: Academic Press; 2012. [Google Scholar] 55. Koci MD, Schultz-Cherry S. Птичьи астровирусы. Avian Pathol. 2002; 31: 213–227. [PubMed] [Google Scholar] 56. Koci MD, Seal BS, Schultz-Cherry S. Разработка диагностического теста RT-PCR для птичьего астровируса. J Virol Methods. 2000. 90: 79–83. [Бесплатная статья PMC] [PubMed] [Google Scholar] 58.Кул Д.А., Холмс И.Х. Нуклеотидная и расчетная аминокислотные последовательности Ty-1 Vp7 ротавируса птиц значительно отличаются от таковых ротавируса Ch-2. Arch Virol. 1993. 129 (1–4): 227–234. [PubMed] [Google Scholar] 59. Lin TL, Loa CC, Tsai SC, Wu CC, Bryan TA, Thacker HL, Hooper T., Schrader D. Характеристика коронавируса индейки у индюшат с острым энтеритом. Vet Microbiol. 2002. 84 (1–2): 179–186. [Бесплатная статья PMC] [PubMed] [Google Scholar] 60. Линь Т.Л., Лоа СС, Ву СС. Полные последовательности 3′-концевой кодирующей области для генов структурных белков коронавируса индейки.Virus Res. 2004. 106 (1): 61–70. [Бесплатная статья PMC] [PubMed] [Google Scholar] 61. Loa CC, Wu CC, Lin TL. Сравнение 3′-концевых кодирующих областей изолятов коронавируса индейки из Индианы, Северной Каролины и Миннесоты со штаммами коронавируса инфекционного бронхита кур. Интервирология. 2006. 49 (4): 230–238. [Бесплатная статья PMC] [PubMed] [Google Scholar] 62. Lojkic I, Bidin M, Bidin Z, Mikec M. Вирусные агенты, связанные с энтеритом домашней птицы в хорватских промышленных стадах индейки. Acta Vet BRNO. 2010; 79: 91–98.[Google Scholar] 63. Маттейнссенс Дж., Мартелла В., Ван Ранст М. Оценка приоритетной статьи: геномная эволюция, межвидовой барьер хозяина, реассортировка и классификация ротавирусов. Будущий Virol. 2010. 5: 385–390. [Google Scholar] 64. Matthijnssens J, Otto PH, Ciarlet M, Desselberger U, Van Ranst M, Johne R. Значения отсечения на основе последовательности VP6 в качестве критерия для демаркации видов ротавирусов. Arch Virol. 2012. 157 (6): 1177–1182. [PubMed] [Google Scholar] 65. Maurel S, Toquin D, Briand FX, Queguiner M, Allee C, Bertin J, Ravillion L, Retaux C, Turblin V, Morvan H, Eterradossi N.Первые полноразмерные последовательности гена S европейских изолятов выявили дальнейшее разнообразие коронавирусов индейки. Avian Pathol. 2011; 40: 179–189. [PubMed] [Google Scholar] 66. Макналти М.С., Аллан Г.М., Маккракен Р.М. Экспериментальное заражение цыплят ротавирусами: клинические и вирусологические данные. Avian Pathol. 1983; 12: 45–54. [PubMed] [Google Scholar] 67. Мор С.К., Абин М., Коста Дж., Дуррани А., Джиндал Н., Гоял С.М., Патнаяк Д.П. Роль астровируса индейки 2 типа в синдроме энтерита домашних птиц. Poult Sci.2011; 90: 2747–2752. [Бесплатная статья PMC] [PubMed] [Google Scholar] 68. Мор С.К., Шарафельдин Т.А., Абин М., Кромм М., Портер Р.Э., Гоял С.М., Патнаяк Д.П. Возникновение кишечных вирусов при синдроме легкой индейки. Avian Pathol. 2013. 42 (5): 497–501. [PubMed] [Google Scholar] 69. Мор С.К., Шарафельдин Т.А., Портер Р.Э., Зиглер А., Патнаяк Д.П., Гоял С.М. Выделение и характеристика реовируса артрита индейки. Птичий Дис. 2013. 57 (1): 97–103. [PubMed] [Google Scholar] 70. Моура-Альварес Дж., Чакон СП, Сканавини Л.С., Нуньес ЛФН, Астольфи-Феррейра К.С., Джонс Р.К., Пиантино Феррейра А.Дж.Кишечные вирусы в стадах бразильских индейок: частота единичных и множественных вирусных инфекций в зависимости от возраста и клинических признаков кишечного заболевания. Poult Sci. 2013; 92: 945–955. [Бесплатная статья PMC] [PubMed] [Google Scholar] 71. Otto PH, Ahmed MU, Hotzel H, Machnowska P, Reetz J, Roth B, Trojnar E, Johne R. Обнаружение птичьих ротавирусов групп A, D, F и G у больных кур и индеек из Европы и Бангладеш. Vet Microbiol. 2012; 156 (1-2): 8-15. [Бесплатная статья PMC] [PubMed] [Google Scholar] 72.Отто П., Либлер-Тенорио Е.М., Эльшнер М., Ритц Дж., Лорен У., Диллер Р. Обнаружение ротавирусов и поражений кишечника у цыплят-бройлеров в стадах с синдромом короткого роста и задержки роста (RSS) Avian Dis. 2006; 50: 411–418. [PubMed] [Google Scholar] 73. Пакпиньо С., Лей Д.Х., Барнс Х.Дж., Вайланкур Дж. П., Гай Дж. С.. Распространенность энтеропатогенной Escherichia coli в естественных случаях синдрома энтерита-смертности домашних птиц. Птичий Дис. 2002. 46 (2): 360–369. [PubMed] [Google Scholar] 74. Palade EA, Demeter Z, Hornyak A, Nemes C, Kisary J, Rusvai M.Высокая распространенность парвовируса индейки в стадах индейки из Венгрии, страдающих синдромами кишечных заболеваний. Птичий Дис. 2011; 55 (3): 468–475. [PubMed] [Google Scholar] 75. Пантин-Джеквуд MJ, Day JM, Jackwood MW, Spackman E. Кишечные вирусы, обнаруженные молекулярными методами в коммерческих стадах кур и индейок в Соединенных Штатах в период с 2005 по 2006 гг. Птичий Дис. 2008. 52: 235–244. [PubMed] [Google Scholar] 76. Пантин-Джеквуд MJ, Spackman E, Day JM. Патогенез астровирусов индейки 2 типа с вариантными генами капсида у 2-дневных цыплят, свободных от специфических патогенов.Avian Pathol. 2008; 37: 193–201. [PubMed] [Google Scholar] 77. Пантин-Джеквуд MJ, Spackman E, Day JM, Rives D. Периодический мониторинг промышленных индеек на предмет энтеросолюбии указывает на постоянное присутствие астровируса и ротавируса на фермах. Птичий Дис. 2007. 51: 674–680. [PubMed] [Google Scholar] 78. Пантин-Джеквуд MJ, Spackman E, Woolcock PR. Молекулярная характеристика и типирование астровирусов кур и индейки, циркулирующих в США: значение для диагностики. Птичий Дис. 2006; 50: 397–404.[PubMed] [Google Scholar] 79. Пантин-Джеквуд MJ, Strother KO, Mundt E, Zsak L, Day JM, Spackman E. Молекулярная характеристика птичьих астровирусов. Arch Virol. 2011; 156: 235–244. [PubMed] [Google Scholar] 80. Куреши М.А., Саиф Ю.М., Хегген-Пи К.Л., Эденс Ф.В., Хавенштейн ГБ. Индукция функциональных дефектов в макрофагах астровирусом индейки, ассоциированным с энтеритом и синдромом смертности домашних птиц. Птичий Дис. 2001. 45 (4): 853–861. [PubMed] [Google Scholar] 81. Рамиг РФ, Ciarlet M, Mertens PPC, Dermody TS. Ротавирус.В: Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA, редакторы. Таксономия вирусов: восьмой отчет международного комитета по таксономии вирусов. Амстердам: Academic Press; 2005. С. 484–496. [Google Scholar] 82. Рейнольдс Д.Л., Саиф Ю.М. Астровирус: причина кишечного заболевания у индюшат. Птичий Дис. 1986; 30: 728–735. [PubMed] [Google Scholar]83. Роббинс К.М. Эпидемиология и диагностика коронавируса в Турции. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс.2013.
84. Родригес-Диас Дж., Рубилар-Абреу Э., Шпицнер М., Хедлунд К.О., Липранди Ф., Свенссон Л. Дизайн мультиплексной вложенной ПЦР для генотипирования NSP4 ротавируса группы А. J Virol Methods. 2008. 149: 240–245. [PubMed] [Google Scholar] 85. Ruff MD, Rosenberger JK. Сопутствующие инфекции реовирусами и кокцидиями у бройлеров. Птичий Дис. 1985. 29 (2): 465–478. [PubMed] [Google Scholar] 86. Шульц-Черри С., Капчинский Д.Р., Симмонс В.М., Кочи М.Д., Браун С., Барнс Х.Дж. Идентификация агента (ов), связанного с синдромом смертности от энтерита домашней птицы: важность тимуса.Птичий Дис. 2000. 44: 256–265. [PubMed] [Google Scholar] 87. Шуман Т., Хотцель Х., Отто П., Джон Р. Доказательства межвидовой передачи и реассортации среди ротавирусов птичьей группы А. Вирусология. 2009. 386 (2): 334–343. [PubMed] [Google Scholar] 88. Продавцы HS, Linnemann EG, Pereira L, Kapczynski DR. Филогенетический анализ гена белка сигма 2 реовирусов индейки. Птичий Дис. 2004. 48 (3): 651–657. [PubMed] [Google Scholar] 89. Продавцы HS, Koci MD, Linnemann E, Kelley LA, Schultz-Cherry S. Разработка диагностического теста мультиплексной полимеразной цепной реакции с обратной транскрипцией, специфичного для астровируса и коронавируса индейки.Птичий Дис. 2004. 48: 531–539. [PubMed] [Google Scholar] 90. Shivaprasad HL, Franca M, Woolcock PR, Nordhausen R, Day JM, Pantin-Jackwood M. Миокардит, связанный с реовирусом у индюшат. Птичий Дис. 2009. 53: 523–532. [PubMed] [Google Scholar] 91. Spackman E, Day JM, Pantin-Jackwood MJ. Сопутствующие инфекции астровируса, реовируса и ротавируса вызывают снижение прибавки в весе у широкогрудых белых цыплят. Птичий Дис. 2010; 54: 16–21. [PubMed] [Google Scholar] 92. Спакман Э., Капчинский Д., Селлерс Х. Мультиплексная полимеразная цепная реакция обратной транскрипции в реальном времени для обнаружения трех вирусов, связанных с комплексом энтерита домашней птицы: астровируса индейки, коронавируса индейки и реовируса индейки.Птичий Дис. 2005; 49: 86–91. [PubMed] [Google Scholar] 93. Спакман Э., Пантин-Джеквуд М., Дэй Дж. М., Селлерс Х. Патогенез реовирусов индейского происхождения у индеек и кур. Avian Pathol. 2005; 34: 291–296. [PubMed] [Google Scholar] 94. Штамм E, Kelley LA, Schultz-Cherry S, Muse SV, Koci MD. Геномный анализ близкородственных астровирусов. J Virol. 2008. 82: 5099–5103. [Бесплатная статья PMC] [PubMed] [Google Scholar] 95. Сугияма М.К., Гото К., Уэмукай Х., Мори Й., Ито Н., Минамото Н. Присоединение и инфицирование к клеткам MA104 ротавирусов птиц требует присутствия сиаловой кислоты на поверхности клетки.J Vet Med Sci. 2004. 66: 461–463. [PubMed] [Google Scholar] 96. Тан Й., Исмаил М.М., Саиф Ю.М. Разработка иммуноферментного анализа с захватом антигена и ОТ-ПЦР для обнаружения астровирусов индейки. Птичий Дис. 2005. 49 (2): 182–188. [PubMed] [Google Scholar] 97. Тан Й., Мурджа А.М., Саиф Ю.М. Молекулярная характеристика гена капсида двух серотипов астровирусов индейки. Птичий Дис. 2005. 49 (4): 514–519. [PubMed] [Google Scholar] 98. MC Teixeira, MC Luvizotto, Ferrari HF, Mendes AR, da Silva SE, Cardoso TC.Обнаружение коронавируса индейки у коммерческих индюшат в Бразилии. Avian Pathol. 2007. 36 (1): 29–33. [PubMed] [Google Scholar] 99. Тейл К.В., Саиф Ю.М. Возрастные инфекции ротавируса, ротавирусоподобного вируса и атипичного ротавируса в стадах индейки. J Clin Microbiol. 1987. 25 (2): 333–337. [Бесплатная статья PMC] [PubMed] [Google Scholar]100. Тилли Б.Дж., Гондер Э., Мейсон С., Уикс Дж. Вспышка коронавируса в Турции в Северной Каролине. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс.2013.
101. Тоффан А., Катания С., Сальвиато А., Де Баттисти С., Васкеллари М., Тосон М., Капуа I, Каттоли Г. Экспериментальное заражение домашних птиц и цесарок генетически отличными птичьими астровирусами. Avian Pathol. 2012; 41: 429–435. [PubMed] [Google Scholar] 102. Trojnar E, Otto P, Roth B, Reetz J, Johne R. Сегменты генома ротавируса группы D имеют консервативные концы группы A, но кодируют специфичные для группы белки. J Virol. 2010; 84: 10254–10265. [Бесплатная статья PMC] [PubMed] [Google Scholar] 103.Trojnar E, Sachsenroder J, Twardziok S, Reetz J, Otto PH, Johne R. Идентификация ротавируса птичьей группы А, содержащего новый ген VP4, близкий родству к генам ротавирусов млекопитающих. J Gen Virol. 2013; 94: 136–142. [PubMed] [Google Scholar] 104. Велаюдхан Б.Т., Шин Х.Д., Лопес В.К., Хупер Т., Халворсон Д.А., Нагараджа К.В. Анализ обратной транскриптазы-полимеразной цепной реакции для диагностики коронавирусной инфекции индейки. J Vet Diagn Invest. 2003. 15 (6): 592–596. [PubMed] [Google Scholar] 105.Woolcock PR, Shivaprasad HL. Электронно-микроскопическая идентификация вирусов, связанных с энтеритом домашней птицы, у индеек, выращенных в Калифорнии в 1993–2003 гг. Птичий Дис. 2008. 52: 209–213. [PubMed] [Google Scholar]106. Вуминг Б. Вспышка коронавируса на северо-западе Арканзаса. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс. 2013.
107. Ю М., Исмаил М. М., Куреши М. А., Дерт Р. Н., Барнс Х. Дж., Саиф Ю. М.. Вирусные агенты, связанные с энтеритом и синдромом смертности домашних птиц: роль небольшого круглого вируса и коронавируса индейки.Птичий Дис. 2000. 44 (2): 297–304. [PubMed] [Google Scholar] 108. Ю М, Тан Й, Го М, Чжан Кью, Саиф Ю. Характеристика небольшого круглого вируса, связанного с синдромом энтерита и смертности цыплят. Птичий Дис. 2000. 44 (3): 600–610. [PubMed] [Google Scholar] 109. Zsak L, Strother KO, Kisary J. Анализ частичной последовательности генома парвовирусов, связанных с кишечным заболеванием у домашней птицы. Avian Pathol. 2008. 37 (4): 435–441. [PubMed] [Google Scholar]Кишечных вирусов энтерита индейки
Вирусная болезнь.2014 июн; 25 (2): 173–185.
, , иНареш Джиндал
Департамент ветеринарного здравоохранения и эпидемиологии, Колледж ветеринарных наук, Университет ветеринарии и зоотехники Лала Ладжпат Рай, Хисар, 125 004 Индия
9004, Индия. К. МорКафедра ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Сент-Пол, Миннесота 55108 США
Сагар М.Гоял
Департамент ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Сент-Пол, Миннесота 55108 США
Департамент ветеринарного здравоохранения и эпидемиологии, Колледж ветеринарных наук, Ветеринарный университет Лала Ладжпат Рай. Науки о животных, Хисар, 125 004 Индия
Департамент ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Сент-Пол, Миннесота 55108 США
Автор, отвечающий за переписку.Поступило 2 октября 2013 г .; Принято 27 января 2014 г.
Эта статья цитируется в других статьях в PMC.Abstract
Здоровье кишечника очень важно для получения максимальной отдачи с точки зрения увеличения веса и яйценоскости. Кишечные заболевания, такие как комплекс энтерита домашней птицы (PEC) у индеек, не позволяют максимально реализовать их производственный потенциал. Был вовлечен ряд вирусов, бактерий и простейших, но первичная этиология окончательно не установлена. Раньше электронная микроскопия использовалась для обнаружения кишечных вирусов, которые определялись исключительно на основе их морфологии.С появлением методов быстрой молекулярной диагностики и секвенирования нуклеиновых кислот нового поколения исследователи добились больших успехов в идентификации и характеристике вирусов, связанных с PEC. Молекулярные методы также помогли нам идентифицировать патогены, которые ранее не были известны. Региональные и национальные исследования выявили наличие нескольких различных кишечных вирусов в PEC, включая ротавирус, астровирус, реовирус и коронавирус, по отдельности или в комбинации.По-прежнему могут существовать неизвестные патогены, которые могут прямо или косвенно играть роль в энтерите у индеек. Этот обзор будет посвящен роли коронавируса, ротавируса, реовируса и астровируса индейки в энтерите индейки.
Ключевые слова: Энтерит, Индейки, Коронавирус, Ротавирус, Реовирус, Астровирус
Введение
На протяжении многих лет у индеек было описано несколько кишечных заболеваний, а именно: коронавирусный энтерит индеек, синдром мальдигестии, синдром ранения и задержки роста у индеек, синдром мальабсорбции у домашней птицы, синдром энтерита и смертности домашних птиц (PEMS), пиковая смертность индеек и вирусный энтерит индейки.Все эти синдромы были включены в комплекс энтерита домашней птицы (PEC), общий термин, обозначающий инфекционные кишечные заболевания молодняка индеек [6]. ПЭК — это экономически важное заболевание молодняка индеек, характеризующееся энтеритом, диареей, умеренной или выраженной депрессией роста, нарушением использования корма, плохой прибавкой веса и в некоторых случаях высокой смертностью, как при ПЕМС [6]. Ряд вирусов (коронавирус, калицивирус, реовирус, астровирус, ротавирус, пикорнавирус, пикобирнавирус, парвовирус и аденовирус), бактерий ( Escherichia coli , Salmonella spp., Clostridia, Campylobacter и Enterococcus ) и простейшие (кокцидии и криптоспоридиумы) участвуют в PEC [6, 18, 47, 73, 105, 109]. Недавно у индюшат Миннесоты были описаны два новых синдрома, а именно «синдром энтерита домашней птицы», или PES, и «синдром легкой индейки», или LTS. ПЭС — это инфекционное кишечное заболевание молодых индеек в возрасте от 1 дня до 7 недель, которое характеризуется диареей, депрессией и летаргией с бледным кишечником и / или чрезмерно жидким содержимым слепой кишки [47].LTS — это проблема индеек товарного возраста, имеющих меньшую массу тела по сравнению с их стандартными породными характеристиками [68]. Мы полагаем, что энтерит у домашних птиц в молодом возрасте создает условия для будущего развития LTS у более старых птиц. Кроме того, мы рассматриваем как PES, так и LTS как часть PEC.
Кишечные вирусы являются частой причиной первичного повреждения желудочно-кишечного тракта (ЖКТ) молодых цыплят, создавая благоприятную среду для роста бактерий и / или простейших и вызывая дальнейшее повреждение кишечника.Возможно, что повреждение кишечника сначала кишечными вирусами, а затем вторичными патогенами может привести к необратимым изменениям в организме хозяина, что может привести к снижению массы тела при продаже. Цель этого обзора — подробно обсудить четыре наиболее важных вируса, вызывающих энтерит индейки, а именно коронавирус, реовирус, ротавирус и астровирус.
Коронавирус
Коронавирусный энтерит индейки — острое заразное заболевание, характеризующееся тупостью, депрессией, диареей и сниженным набором массы тела.Заболевание было впервые описано в 1940-х годах в штате Вашингтон, США. Возбудитель не был идентифицирован, и заболевание было названо «грязевой лихорадкой». Однако после выявления в этих случаях коронавируса индейки (TCoV) название было изменено на «коронавирусный энтерит индеек». В 1970-х годах эта болезнь имела самые разрушительные последствия для индеек из Миннесоты. Были предприняты шаги по искоренению болезни, которые привели к элиминации этого вируса из Миннесоты. В настоящее время TCoV спорадически регистрируется из регионов Северной Америки и других стран, где выращивают индейку.TCoV также связан с PEMS, острым высококонтагиозным кишечным заболеванием, характеризующимся депрессией, анорексией, диареей и высокой смертностью индеек.
Геном
Коронавирусы представляют собой плеоморфные частицы с оболочкой, примерно сферической формы, диаметром от 50 до 200 нм. Согласно последнему предложению Международного комитета по таксономии вирусов (ICTV), семейство Coronaviridae включает два подсемейства: Coronavirinae и Torovirinae. Coronavirinae состоит из трех родов: Alphacoronavirus , Betacoronavirus и Gammacoronavirus [54], из которых последний включает коронавирусы птиц. Геном коронавируса состоит из одноцепочечной РНК с положительным смыслом. Полноразмерный геном состоит из 27 632 нуклеотидов плюс 3 ‘поли (А) хвост. Есть две открытые рамки считывания (ORF), ORF 1a и 1b, которые находятся в первых двух третях генома с девятью дополнительными ORF ниже по течению.Область между генами белков мембраны (M) и нуклеокапсида (N) содержит три потенциальных малых ORF: (1) ORF-X, (2) ранее не охарактеризованную ORF со связанной предполагаемой TRS в гене M (очевидно, общей для всей группы III coronaviruses) и (3) ранее описанные ORF 5a и 5b. TCoV не содержит гена гемагглютинин-эстеразы [29].
Сообщалось о вариабельности в различных генных сегментах TCoV. Сравнение последовательностей полной 3′-концевой области, кодирующей структурный белок четырех TCoV, с соответствующими последовательностями вируса инфекционного бронхита (IBV) показало, что TCoV, вероятно, имеет то же происхождение, что и IBV, и приобретенные последовательности S-гена для тропизма кишечника индейки во время процесс эволюции [61].Lin et al. [60] сообщили, что последовательность S-белка TCoV имеет только 33,8–33,9% идентичности с IBV, хотя оба вируса сгруппированы в одну и ту же геномную линию. Полноразмерные последовательности гена spike (S) французского происхождения TCoV обладали 98% идентичностью как на уровне нуклеотидов, так и на уровне аминокислот [65]. Однако эти вирусы показали 65 и 60% сходства с североамериканскими TCoV и 50 и 37% сходства с IBV на уровне нуклеотидов и аминокислот, соответственно.
Заболеваемость и распространение
В отличие от обычно неопределенных синдромов вирусных кишечных заболеваний, TCoV-энтерит можно напрямую отнести к инфекции, вызванной TCoV [35, 38, 41].TCoV распространен во всем мире и был обнаружен у пораженных энтеритом индеек из Северной Америки, Южной Америки и Европы [34, 59, 65, 70, 98]. Было обнаружено, что 37% кишечных проб от больных стад индейки в Европе оказались положительными на TCoV [65]. В недавнем исследовании, проведенном в Бразилии, большинство образцов кишечника, взятых как у пораженных энтеритом, так и у здоровых индеек, оказались положительными на TCoV [70]. В этом исследовании 55,3% TCoV-положительных образцов также были положительными на другие кишечные вирусы.Недавно были зарегистрированы вспышки коронавирусного энтерита из Арканзаса и Северной Каролины в США с тяжелой диареей и смертностью [19, 83, 100, 106], что указывает на важность постоянного эпидемиологического надзора за этим вирусом. TCoV, по-видимому, отсутствует в синдромах ПЭК без ПЭМС [44, 45, 62, 68, 75, 105]. Например, коронавирус не был обнаружен, когда в Калифорнии были протестированы 2400 образцов от домашних птиц с клиническими признаками энтерита [105]. Аналогичным образом, в исследовании 33 стай индеек, пораженных энтеритом, в США, Pantin-Jackwood et al.[75] не нашли TCoV. Мы также не обнаружили TCoV в случаях PES и LTS [44, 45, 68].
TCoV может поражать птиц любой возрастной группы, но вызывает высокую смертность молодых индюшат в возрасте 1–4 недель. Смертность колеблется от менее 10 до 50%. Индюки, по-видимому, являются единственным естественным хозяином TCoV; при экспериментальной инокуляции TCoV вызывал заболевание у домашних птиц, но не у цыплят [42]. Репликация TCoV происходит в энтероцитах, выстилающих апикальную часть ворсинок кишечника. Разрушение эпителия ворсинок кишечника приводит к диарее, нарушению пищеварения и мальабсорбции, что может привести к снижению прибавки в весе у индюшат.
Передача
Вирус присутствует в основном в кишечнике и фекалиях пораженных домашних птиц, следовательно, передача вируса осуществляется фекальным оральным путем. Прямая передача происходит при прямом контакте с инфицированными птицами, тогда как косвенная передача происходит механическим путем через рабочих и / или зараженное оборудование. Однако в экспериментальном исследовании антиген TCoV также может быть обнаружен в придаточных пазухах носа и слезной добавочной железе (железе Хардера) инфицированных цыплят в течение 14 дней после инокуляции [33].Вирус выделяется с фекалиями индеек в течение многих недель после выздоровления от клинического заболевания. Большинство случаев заражения происходит летом (май – август), иногда осенью. TCoV может сохраняться в течение длительного периода времени и может легко передаваться между птицефабриками в прохладной окружающей среде.
Клинические признаки и патология
Индюки, пораженные TCoV, сбиваются в кучу и демонстрируют взъерошенные перья, снижение потребления корма и воды, влажный помет и потерю живой массы.У пожилых птиц может наблюдаться депрессия, диарея и задержка роста [34]. Помет водянистый и пенистый, может содержать слизь, от зеленого до коричневого цвета. Клинические признаки могут сохраняться до 2 недель, а восстановление веса, если оно будет достигнуто, может занять несколько недель. У несушек наблюдается снижение яйценоскости [3] из-за аномальной пигментации яиц. Птицы в возрасте от 1 до 4 недель, у которых наблюдаются диарея, обезвоживание и задержка роста на 40% или более, считаются страдающими PEMS. Существует две формы PEMS: (1) тяжелая форма i.е. рост нравственности индеек (SMT), который определяется как смертность, равная или превышающая 1% в течение 3 дней подряд или 9% в возрасте от 8 до 28 дней; и (2) легкая форма, которая определяется как смертность более 2%, но менее 9% в течение того же периода времени, но не равная или не превышающая 1% в течение трех дней подряд. В экспериментальном исследовании, как заболеваемость, так и смертность регистрировались у домашних птиц, экспериментально инокулированных TCoV в возрасте 2 дней, однако птицы, инфицированные в возрасте 28 дней, имели значительную задержку роста, но не демонстрировали смертности [32].
TCoV может действовать синергетически с другими патогенами, увеличивая тяжесть заболевания, или может предрасполагать птиц к вторичным инфекциям. У цыплят, инокулированных только TCoV, развилась умеренная депрессия роста без гибели, а у свиней, инокулированных только энтеропатогенной Escherichia coli (EPEC) , клиническое заболевание не развилось. Однако инокуляция цыплят TCoV и EPEC одновременно вызвала повышение смертности, подавление роста и прикрепление / стирание повреждений [38].
У пораженных птиц наблюдается задержка роста и обезвоживание. Кишечник заметно увеличен и заполнен рыхлым желтым содержимым. Стенки кишечника вялые и бледные. Caeca также вздуты с пенистым и водянистым содержимым. Крупные изменения, такие как атрофия поджелудочной железы, селезенки и бурсы Фабрициуса, также могут наблюдаться наряду с расширением желчного пузыря и мочеточников. У инфицированных кур наблюдаются более выраженные поражения яичников с деформированными, плохо развитыми яичниками и скоплением казеозного экссудата в брюшине.Также наблюдаются спайки брюшины яйцевода.
В зависимости от тяжести инфекции наблюдается от легкого мультифокального до тяжелого энтерита с атрофией ворсинок. Наблюдается слущивание эпителиальных клеток с застоем и обширной инфильтрацией лимфоцитов и гетерофилов в собственную пластинку ворсинок. Кроме того, также может наблюдаться истощение фолликулярных лимфоидов и атрофия коркового вещества тимуса.
Диагноз
Клинические признаки, такие как диарея, депрессия, скученность и задержка роста, указывают на кишечную инфекцию.Высокая смертность молодняка может указывать на ПЕМС.
Выделение вируса, электронная микроскопия (ЭМ), обнаружение вирусной РНК и серология могут помочь в диагностике. Вирус можно выращивать в зародышах индейки или куриных яйцах, но его трудно выращивать в клеточных культурах. ЭМ с отрицательным контрастом может выявить вирусные частицы размером ~ 50–200 нм с типичной морфологией в форме короны в содержимом кишечника. Прямые и непрямые флуоресцентные тесты на антитела и ELISA использовались для обнаружения антигена TCoV и антител соответственно [30, 31, 37].О высокой распространенности TCoV сообщалось от заводчиков (71,1%) и мясных индеек (56,7%) в Онтарио, Канада [31]. В Арканзасе 64,2% образцов сыворотки дали положительный результат на антитела [30].
Доступны молекулярные тесты для обнаружения и дифференциации TCoV от других кишечных вирусов. Доступны различные тесты полимеразной цепной реакции с обратной транскрипцией (RT-PCR), нацеленные на 3′-нетранслируемую область (UTR), S, N и / или M ген TCoV [11, 12, 16, 89, 92, 104]. Также была разработана ОТ-ПЦР в реальном времени, которая является наиболее чувствительным и специфическим тестом, с помощью которого можно количественно определить вирусную нагрузку в тканях индейки, а также в образцах фекалий [15].Прямая иммуногистохимия была рекомендована в качестве альтернативы ОТ-ПЦР для обнаружения TCoV в полевых образцах [14].
Профилактика и контроль
Специального лечения или вакцины нет. Биобезопасность, при которой особое внимание уделяется управлению подстилкой и надлежащей утилизации мертвых птиц и использованного подстилки, похоже, единственный способ предотвратить распространение инфекции. Кроме того, общие меры, такие как уменьшение влажности подстилки, использование антибиотиков для борьбы с вторичными бактериальными инфекциями и улучшение гигиены, могут минимизировать последствия энтерита TCoV.
Реовирус
Птичий реовирус был впервые выделен от кур, страдающих хроническим респираторным заболеванием, и получил название возбудитель Фейи-Кроули (FC). Позже возбудитель FC был охарактеризован как реовирус. Однако у кур птичий реовирус вызывает множество синдромов, включая вирусный артрит / теносиновит. Недавно реовирусы были выделены от артрита / тендосиновита индейки и получили название TARV (реовирус артрита индейки) [69]. Реовирусы в основном бывают двух типов: фузогенные и нефузогенные.Слитые вирусы обладают способностью вызывать слияние инфицированных клеток, что приводит к образованию многоядерных синцитий, в то время как нефузогенные вирусы не вызывают слияния инфицированных клеток. К нефузогенным типам относятся, в основном, реовирусы млекопитающих (MRV), тогда как слитные реовирусы поражают млекопитающих, птиц и рептилий.
Геном
Реовирус принадлежит к роду Orthoreovirus в семействе Orthoreoviridae. Вирус без оболочки, имеет икосаэдрическую симметрию и размер частиц 70–80 нм.Он содержит геном двухцепочечной (ds) РНК из десяти сегментов, которые делятся на три класса, называемые большими (L), средними (M) и малыми (S), в зависимости от их характера миграции при электрофорезе в полиакриламидном геле. Гены L и M далее подразделяются на три сегмента каждый (L1, L2, L3 и M1, M2, M3, соответственно), в то время как ген S имеет четыре сегмента (S1, S2, S3, S4) [7].
Геном реовируса имеет 12 открытых рамок считывания, которые кодируют восемь структурных и четыре неструктурных белка.Структурные белки являются важной частью вирионов потомства, в то время как неструктурные белки не присутствуют в зрелом вирионе и экспрессируются только в инфицированных клетках. Структурными белками, кодируемыми генами L, M и S, являются лямбда (λ), мю (μ) и сигма (σ) соответственно. Три структурных белка λA, λB и λC кодируются сегментами L гена L1, L2 и L3 соответственно. Сегменты M1 и M2 кодируют два структурных белка (μA и μB) соответственно, в то время как сегмент M3 кодирует неструктурный белок (μNS).Три σ-белка σC, σA и σB кодируются сегментами S1, S2 и S3 соответственно, тогда как сегмент S4 кодирует неструктурный белок σNS. Сегмент S1 кодирует два дополнительных неструктурных белка, а именно p10 и p17 [7, 10].
На основании анализа последовательностей различных генов сообщалось, что реовирусы индейки отличаются от реовирусов кур [20, 44, 45, 52, 88]. Например, сходство нуклеотидной и аминокислотной последовательности сегмента S3, кодирующего сигма-B-белок реовируса индейки (TRV), было обнаружено примерно 61 и 78-80%, соответственно, по сравнению с изолятами куриного реовируса [88].Day et al. [20] сообщили, что белок сигма C гена S1 TRV имеет 57% аминокислотную идентичность с таковой у куриного реовируса (эталонный штамм S1133), тогда как белки p10 и p17 имеют 72 и 61% идентичности, соответственно, с соответствующими белками S1133.
Белок сигма С реовируса млекопитающих и слитый реовирус Нельсона Бэй на 25–28% имеют аминокислотную идентичность с белком сигма С TRV. Ранее мы сообщали, что сходство последовательностей гена S4 на уровне нуклеотидов TRVs было 71.1–74,2% по сравнению с реовирусами куриного происхождения [45]. Эти исследования показывают, что TRV образуют отдельную группу по сравнению с куриными и другими изолятами птичьего реовируса. В результате TRV потенциально можно рассматривать как отдельный вид вируса в подгруппе 2 рода Orthoreovirus . Эта точка зрения дополнительно подтверждается экспериментальными исследованиями, в которых TRV не смогли вызвать признаки заболевания у цыплят, хотя они могут реплицироваться на низких уровнях в кишечном тракте цыплят.Более того, высокопатогенный штамм реовируса куриного происхождения (ARV-1733), вызывающий высокую смертность цыплят, не вызывал клинических признаков у промышленных индеек и имел лишь легкие клинические признаки у индюков, свободных от специфических патогенов [93].
Заболеваемость и распространение
Около 85–90% изолятов реовируса не являются патогенными. Патогенные штаммы реовирусов были связаны с различными заболеваниями кур и индеек. У индеек этот вирус был обнаружен при нескольких различных кишечных заболеваниях, а именно LTS, PEC, PES, миокардите, а недавно и при артрите / теносиновите [6, 28, 39, 44, 47, 68, 69, 90, 91].При исследовании 33 стад индюшат 46% стад оказались положительными на реовирусы [75]. Jindal et al. [44] проверили содержимое кишечника 43 стад, пораженных PES, в 2007–2008 гг. И сообщили о наличии реовируса в 40% стад. Энтерит, вызванный реовирусом, в основном встречается у молодняка индейки в возрасте 1–7 недель, а заболеваемость выше у птиц в возрасте 1–3 недель, которая уменьшается с увеличением возраста.
Реовирусы обнаружены также у практически здоровых кур и индеек [45, 68, 75].При исследовании пяти здоровых родительских стад индюшат 10,4% из 193 протестированных образцов (в возрасте от 1 до 9 недель) были положительными на реовирус [45]. Тестирование образцов кишечника 33 промышленных стад индейки из всех регионов США в течение 2005 и 2006 гг. Выявило присутствие реовируса в 45,5% стад [75].
Передача
Горизонтальная передача фекально-оральным путем является обычным явлением. Из-за стабильности вируса в окружающей среде также может происходить механическая передача.Различные штаммы реовируса различаются по способности распространяться по горизонтали. Подозревается, но не доказано, вертикальная передача кишечной реовирусной инфекции.
Клинические признаки и патология
Пораженные птицы страдают диареей, депрессией, взъерошенными перьями, а также сниженным набором веса [93] и смертностью. Инфекция АРВ может также предрасполагать птиц к вторичным осложнениям или действовать синергетически вместе с другими инфекциями. Сообщалось, что АРВ-инфекция может усиливать кокцидиальную инфекцию у цыплят-бройлеров [85].Также существует возможность синергического действия TRV с другими патогенами у индеек.
В большинстве случаев содержимое кишечника пенистое и водянистое. В некоторых случаях наблюдается атрофия бурсы Фабрициуса [22, 93]. Поражения кишечника состоят из легкой гиперплазии крипт из-за инфильтрации лимфоцитов, гетерофилов и эозинофилов в собственную пластинку и подслизистую оболочку через 11–14 дней после заражения. В сумке Фабрициуса наблюдается умеренное или тяжелое истощение фолликулярных лимфоцитов [22, 93].Помимо атрофии бурсальной сумки, может наблюдаться лимфоидное истощение от легкой до умеренной степени в селезенке и лимфоцитарная инфильтрация в печени, поджелудочной железе, сердце и преджелудках.
Диагноз
Диарея в стаде является признаком кишечной инфекции. Трудно точно определить вирус / возбудитель только на основе диареи, и для подтверждения необходимы лабораторные тесты. Электронная микроскопия может выявить присутствие вирусных частиц размером ~ 70–80 нм в кишечнике инфицированных птиц.Вирус хорошо растет в курином яйце с эмбрионом через желточный мешок или хориоаллантоисную мембрану, а также был выделен в первичных клетках печени куриного эмбриона (CEL), клетках почек куриного эмбриона (CEK) и клетках фибросаркомы японского перепела (QT-35). Из-за фузогенной природы TRV в инокулированных культурах клеток наблюдается характерный цитопатический эффект (ЦПЭ). Для обнаружения антител против реовируса цыплят коммерчески доступны наборы ELISA, но в настоящее время таких специфических наборов для реовируса индейки нет.Недавно мы разработали тест ELISA для обнаружения антител против реовируса индейки. В этом тесте мы клонировали и экспрессировали белок сигма C S1 и использовали его в качестве антигена для обнаружения антител против TRV (неопубликованные данные Mor et al.).
Доступно несколько тестов ОТ-ПЦР, нацеленных на различные генные сегменты реовируса [4, 20, 44, 45, 52, 88]. ОТ-ПЦР в реальном времени — это наиболее чувствительный и специфический тест, с помощью которого можно количественно определить вирусную нагрузку в клинических образцах. Был разработан надежный, сверхчувствительный и точный количественный анализ на АРВ с помощью Light Cycler SYBR Green на основе ОТ-ПЦР в реальном времени, который может обнаруживать 39 копий / мкл геномной РНК АРВ [53].Недавно в нашей лаборатории была разработана высокочувствительная и специфическая ОТ-ПЦР в реальном времени для обнаружения TRV. Мы разработали праймеры и зонды из сегмента генома S4. Тест специфичен для TRV (энтерических вирусов индейки и реовирусов артрита) и может обнаруживать до 10 копий транскрибированной in vitro стандартной вирусной РНК на реакцию (Mor et al. Неопубликованные данные).
Профилактика и контроль
Эффективного лечения реовирусных инфекций не существует. Вирус является повсеместным по своей природе, и из-за его высокой стабильности в окружающей среде трудно избавить птиц и фермы от реовируса.В настоящее время вакцины против реовируса индейки нет, и поэтому биозащита остается единственным способом борьбы с инфекцией.
Ротавирус
Ротавирусы являются важными патогенами, которые вызывают неонатальную диарею у людей, а также у многих животных и птиц. Этот вирус также был обнаружен у коммерческих цыплят-бройлеров, страдающих синдромом короткого роста и задержки роста [72].
Геном
Ротавирусы — это РНК-вирусы, принадлежащие к роду Rotavirus и семейству Reoviridae .Геном ротавируса состоит из 11 сегментов двухцепочечной РНК; РНК окружена тройным слоем икосаэдрического белкового капсида. Каждый сегмент кодирует как минимум один белок. Внешний белковый слой состоит из двух основных нейтрализующих антигенов — вирусного белка 4 (VP4) и вирусного белка 7 (VP7). Вирусный белок 6 (VP6) второго слоя капсида называется групповым антигеном, который используется для обнаружения и классификации ротавирусов на отдельные группы. Кроме того, по крайней мере пять неструктурных белков кодируются геномом ротавируса.
Основываясь на антигенных свойствах VP6, ICTV разделил ротавирусы на пять серологических видов (A – E) и два дополнительных вида (F и G) [81], которые также называются группами ротавирусов. Известно, что группы A, B и C инфицируют людей и животных, а группы D, E, F и G заражают только животных, в основном птиц [50, 63, 102]. Недавно было сообщено о потенциально новом виде RV, ротавирусе H [ 64 ]. Эти авторы утверждают, что значение отсечения аминокислотной последовательности 53% позволяет дифференцировать различные виды ротавирусов.
В отличие от ротавирусов птиц группы A, ротавирусы группы D не могут размножаться в клетках MA104 [ 25 ] и в основном идентифицируются по схеме электрофоретической миграции 5: 2: 2: 2 их сегментов генома, что значительно отличается от паттерна 5: 1: 3: 2 ротавирусов птичьей группы А. Однако классификация штамма ротавируса только на основе его профиля миграции РНК является проблематичной из-за возможного возникновения перестройки генома. В отличие от ротавирусов группы А, которые преимущественно инфицируют клетки двенадцатиперстной кишки, вирусы группы D имеют склонность к тощей и подвздошной кишке.
Ротавирус индейского происхождения отличается от ротавируса куриного происхождения [58]. Например, сообщалось, что гомология аминокислотной последовательности гена VP7 штамма Ty-1 (индейка) и гена Ch-2 VP7 птиц составляет только 70% [58]. Вариабельные эпитопные области A, B и C Ty-1 были уникальными по сравнению с таковыми у Ch-2 и других штаммов, представляющих серотипы 14G. Низкая гомология (53%) областей A и C Ty-1 и Ch-2 предполагает, что штамм Ty-1 может иметь другой серотип, чем эталонный штамм G7 Ch-2.Сравнение последовательностей различных генов (VP4, VP6, VP7, NSP5) ротавирусов индейки и кур показало, что среди ротавирусов птичьей группы А может происходить межвидовая передача и реассортация [87].
Заболеваемость и распространение
Ротавирусы распространены по всему миру и были зарегистрированы как у здоровых, так и у пораженных энтеритом индеек [44, 45, 69, 71, 75, 77, 99]. Периодический мониторинг товарных стад индейки выявил циркуляцию четырех групп (на основе анализа последовательности гена NSP4) ротавирусов.Среди четырех групп ротавирусов было 96,1–97,5 и 97,5–99,55% сходства на уровне нуклеотидов и аминокислот соответственно [77]. Ротавирусы присутствовали до размещения, а также после размещения до 12-недельного возраста [77]. В другом исследовании, в котором участвовали 33 стада индейки и 43 стада кур, ротавирусы были обнаружены в 46% стадах цыплят и 70% стадах индейки [75]. Мы также сообщали о присутствии ротавирусов в практически здоровых стадах производителей индейки с максимальной встречаемостью до 5-недельного возраста [45].В другом исследовании 93% стад, пораженных PES, были положительными на ротавирусы [44]. В контрольном исследовании Mor et al. [68] сообщили, что шесть (7,5%) пулов образцов фекалий были положительными на ротавирус в стадах, пораженных LTS, в то время как 13 (33%) пулов образцов были положительными из стада без LTS.
Выявление ротавирусов выше у молодых цыплят, и уровень обнаружения снижается с возрастом. При наблюдении PES и LTS максимальное количество цыплят было положительным на ротавирус в возрасте 2–5 недель с последующим уменьшением количества положительных образцов в возрасте 8–9 недель [44, 45, 68].Было показано, что праймеры на основе высококонсервативной области генома ротавируса (в частности, неструктурных белков NSP3 или NSP4) подходят для идентификации ротавируса в различных клинических образцах, образцах животных или окружающей среды с помощью обычных или в реальном времени RT- ПЦР [51, 84]. В исследовании, проведенном в Европе, ротавирусы кур и индеек были идентифицированы с использованием двух разных методов: электрофореза в полиакриламидном геле с ОТ-ПЦР и анализов ОТ-ПЦР в реальном времени, специфичных для ротавирусов групп A и D [71].Эти результаты показывают, что необходимы дальнейшие исследования для точного определения типов ротавирусов, циркулирующих в стадах индейки в различных географических точках.
Передача
Фекальный оральный путь является основным путем горизонтальной передачи. Вирус может выжить в окружающей среде в течение длительного времени, и, следовательно, механическая передача может играть роль в его передаче. Есть подозрение на вертикальную передачу ротавируса, но это пока не подтверждено. Например, Pantin-Jackwood et al.[77] обнаружили ротавирусы в 50% проб, исследованных до помещения птицы. Основываясь на этой информации, авторы предположили, что ротавирус может передаваться вертикально. Мы также обнаружили ротавирусы у 2-дневных коммерческих и родительских домашних птиц [45, 47], что указывает на возможность вертикальной передачи ротавируса.
Также описана межвидовая передача ротавируса [87, 103]. В одном исследовании ротавирусы крупного рогатого скота группы А (n = 6) были обнаружены в кишечном содержимом индюков [2].Электроферотипы показали картину миграции, идентичную вирусу диареи телят в Небраске, а полная филогения гена NSP4 показала, что все шесть штаммов сегрегированы по генотипу E2, что подтверждает их принадлежность к ротавирусу крупного рогатого скота. Ротавирусы птиц не обнаружены у людей, и ротавирусы человека не содержат последовательностей ротавирусов птиц [87], что указывает на низкую вероятность заражения человека ротавирусами птиц.
Клинические признаки и патология
Тупость, депрессия, водянистый помет, потеря аппетита, склеивание вентиляционных отверстий и снижение прибавки в весе — постоянные признаки у пораженных птиц.Смертность может присутствовать или отсутствовать, а заболеваемость может варьироваться. Ротавирусная инфекция кур-несушек может привести к снижению яйценоскости.
В большинстве случаев наблюдается пенистое и водянистое кишечное содержимое. Может наблюдаться бледность кишечного тракта и вздутие слепой кишки с пенистым или непенистым жидким содержимым. Другими крупными патологическими проявлениями являются обезвоживание, задержка роста и пастообразные выделения. Также отмечаются мультифокальные и поверхностные эрозии двенадцатиперстной и тощей кишки. Наиболее частыми микроскопическими изменениями являются гиперцеллюлярность собственной пластинки из-за инфильтрации гетерофилов, эозинофилов и мононуклеарных клеток, а также атрофия ворсинок.Сканирующая электронная микроскопия показывает шероховатость поверхности ворсинок, нарушение нормальной морфологии ворсинок и потерю микроворсинок в клетках, расположенных на кончиках ворсинок. У цыплят разные штаммы ротавирусов могут иметь разные предпочтительные сайты репликации. Например, McNulty et al. [66] сообщили, что у экспериментально инфицированных кур ротавирус группы A имел сродство к двенадцатиперстной кишке, тогда как ротавирус группы D предпочитал тощую кишку и подвздошную кишку.
Диагноз
Диарея, обезвоживание и снижение веса стада свидетельствуют о кишечной инфекции.ЭМ кишечного содержимого и фекалий выявляет присутствие вирусных частиц размером ~ 70 нм (приблизительно 50–70 нм) с характерным «колесом» внешнего вида. Несмотря на то, что вирус трудно изолировать, он все же растет в клетках MA104 при обработке трипсином. Тест коагглютинации использовался для обнаружения ротавирусов у индеек и оказался простым и быстрым скрининговым тестом. Электрофорез в полиакриламидном геле обычно используется для определения характерной картины миграции 11 сегментов генома ротавируса и почти так же чувствителен, как ЕМ.
Для обнаружения вируса доступно несколько ОТ-ПЦР и ОТ-ПЦР в реальном времени, нацеленных на различные сегменты генома ротавируса [8, 21, 77]. Недавно было сообщено о высокочувствительной и специфической одностадийной ОТ-ПЦР в реальном времени, нацеленной на неструктурный белок 4 (NSP4), с внутренней системой контроля для обнаружения ротавирусов индейки [1].
Профилактика и борьба
Специальных мер борьбы с ротавирусными инфекциями не существует. Получение цыплят из источников, свободных от ротавирусов, и принятие надлежащих мер биобезопасности — это способы минимизировать заражение ротавирусом в стаде.
Астровирус
Астровирус индейки (TAstV) был впервые обнаружен у индюшат в возрасте 6–11 дней с диареей и повышенной смертностью в Великобритании. В США TAstV был обнаружен у индюшат и позже был назван TAstV-1. Впоследствии TAstV, связанный с PEMS, был выделен и охарактеризован. Этот вирус генетически и иммунологически отличался от ранее описанного изолята США, TAstV-1, и был назван TAstV-2 [56, 86]. Помимо TAstV-1 и TAstV-2, астровирус куриного происхождения i.е. вирус птичьего нефрита (ВНВ) обнаружен также у индеек [26, 75]. ANV сначала был описан как небольшой круглый вирус, затем как пикорнавирус, а позже был определен как астровирус [40]. Таким образом, у индеек обнаружено три типа астровирусов (TAstV-1, TAstV-2 и ANV). Однако данные о последовательностях предполагают, что в природе может присутствовать гораздо больше типов [94]. В 1980-х годах все обнаруженные астровирусы были строго ограничены желудочно-кишечным трактом домашних птиц, и не было никаких признаков системного заболевания.Однако TAstV также был обнаружен в тимусе и сумке, что указывает на возможное влияние на иммунную систему [86]. Сообщалось также об обнаружении астровирусов у кур и индюшат, страдающих кишечными и / или опорно-двигательными расстройствами (теносиновит) [26, 82].
Геном
Астровирусы — это РНК-вирусы без оболочки диаметром 25–35 нм, принадлежащие к роду Avastrovirus семейства Astroviridae . Геном астровируса состоит из 6,8–7 РНК.9 kb и имеет 5′-UTR, за которым следуют три ORF, 3′-UTR и поли-A-хвост. ORF-1a кодирует сериновую протеазу и другие неструктурные белки, ORF-1b для РНК-зависимой РНК-полимеразы и ORF-2 для капсидного белка. Общая длина геномов трех астровирусов, а именно. TAstV-1 (7003 нт), TAstV-2 (7325 нт) и ANV (6927 нт) варьируются; эта длина не включает хвост поли-А [55].
Большинство исследований по обнаружению и характеристике TAstV было проведено молекулярными методами с использованием праймеров, специфичных для гена капсида или полимеразы.Ряд исследований показал вариации на уровне нуклеотидов и аминокислот в гене капсида и / или гене полимеразы TAstVs [13, 44–46, 78, 79, 97]. Сообщается о высоком уровне генетической изменчивости среди астровирусов индейки из стад индейки в США [78]. Идентичность нуклеотидной последовательности в гене капсида, связанная с серотипом и вирусным патогенезом, была всего лишь 69%, но ген полимеразы был более консервативным с 86–99% идентичностью нуклеотидов. Недавно филогенетический анализ белка-предшественника капсида выявил обширные вариации в аминокислотных последовательностях TAstV-2 (81.5–100%), но не в TAstV-1 (96,2–100%) [79]. Мы также обнаружили вариации на уровне аминокислот в гене капсида TAstV-2 [44, 46]. Эти изменения предполагают, что в природе могут существовать различные серотипы астровируса индейки. Для лучшего понимания серотипов TAstV, циркулирующих у индеек, необходимы исследования полных последовательностей генов капсида из явно здоровых и пораженных PEC стад из разных географических регионов. Изменения гена капсида из-за мутаций или рекомбинации могут влиять на антигенность и патогенность вирусов и, следовательно, иметь практическое значение для методов обнаружения вирусов, эпидемиологических исследований и разработки потенциальных вакцин против астровирусных инфекций.
Заболеваемость и распространение
Был проведен ряд исследований для выявления присутствия TAstV в стадах индейки с использованием различных методов. Ранее вирусы в образцах кишечника выявлялись с помощью прямой или иммунной ЭМ. Однако в большинстве недавних исследований используются молекулярные инструменты, такие как ОТ-ПЦР и секвенирование, для обнаружения различных астровирусов [36, 56, 57, 108]. В ходе исследования восьми промышленных предприятий по выращиванию индейки и одного исследовательского подразделения в США, 89,5% из 96 образцов содержимого кишечника были положительными на астровирус с помощью ОТ-ПЦР [77].В исследовании, проведенном в Миннесоте, 84% из 43 стай индеек, пораженных PES, оказались положительными на TAstV-2 с помощью ОТ-ПЦР [44]. Эти образцы не тестировались на наличие TAstV-1 и ANV, поэтому нельзя исключать присутствие TAstV-1 и ANV в этих образцах. Ряд других исследований также выявил высокую распространенность TAstV в стадах индейки в США [75, 77, 78].
Исследования распространенности TAstV также проводились в других странах, где выращивают индюшат [17, 27, 62, 70, 74]. Да Силва и др.[17] сообщили о TAsV у молодых цыплят, пораженных PEC в Бразилии. В другом исследовании, проведенном в Бразилии, TAsV-1, TAstV-2 и ANV были идентифицированы в 64,5, 44,7 и 35,5% образцов кишечника соответственно [70]. Из них TAstV-1 и TAstV-2 были более распространены в стадах индеек, пораженных энтеритом. Domanska-Blicharz et al. [27] сообщили, что TAstV-2 является наиболее распространенным вирусом в Польше; 30 из 77 стад индейки (38,9%) были положительными. TAstV-1 был обнаружен только в девяти стадах, тогда как ANV — в одном стаде. Эти три вируса были обнаружены либо как одиночные, либо как смешанные инфекции.Тестирование 23 образцов кишечника от откормленных индюков с клиническими признаками энтерита выявило присутствие TAstV-2 в 17 образцах в Хорватии [62].
TAstV были обнаружены не только в стадах индюшат, пораженных энтеритом, но также и в практически здоровых стадах индюшат [45, 68, 78]. Jindal et al. [45] протестировали 193 объединенных образца фекалий практически здоровых индюшат-производителей с помощью ОТ-ПЦР и обнаружили, что 47,2% оказались положительными на TAstV-2.
Передача
Передача астровирусов фекально-оральным путем.Здоровые птицы могут подвергаться воздействию астровируса либо при прямом контакте с инфицированными фекалиями, либо с материалами, загрязненными фекалиями. Считается, что астровирусные инфекции видоспецифичны, но недавние сообщения указывают на межвидовую передачу [23, 101]. Так, TAstV-2 и ANV были обнаружены у уток ( Anas platyrhynchos domesticus ) [9]. Межвидовая передача и адаптация вируса к новым хозяевам или совместное инфицирование одного и того же хозяина разными астровирусами может привести к появлению новых астровирусов, которые инфицируют животных или обладают зоонозным потенциалом [24].
Клинические данные и патология
Как и при рота- и реовирусных инфекциях, астровирусы также вызывают диарею, обезвоживание и задержку роста. Сообщалось о подобных наблюдениях диареи и задержки роста у экспериментальных цыплят, инокулированных материалом, содержащим TAstV [48, 49, 76]. Были предприняты экспериментальные исследования, чтобы установить тип клинического заболевания, вызываемого TAstV, имеющими вариантные капсиды, или те, которые различались по источнику происхождения. Например, Pantin-Jackwood et al.[76] изучали патогенность трех различных вирусов TAstV-2 у индюков, свободных от специфических патогенов, и обнаружили, что вирусы TAstV-2 с вариантными капсидами вызывают подобное кишечное заболевание у молодых индеек. Mor et al. [67] обнаружили, что TAstV-2 от птиц PES был более патогенным, чем TAstV-2 от практически здоровых домашних птиц. Астровирус может потенциально нарушать иммунный ответ, тем самым повышая восприимчивость индеек к вторичным бактериальным инфекциям [80].
Обезвоживание, вздутие кишечника, наполненное водянистым содержимым и непереваренной пищей, и расширенная слепая кишка с пенистым содержимым — это результаты вскрытия.Может возникнуть атрофия бурсальной сумки от легкой до умеренной, а также снижение тонуса кишечника. Гистологическими находками являются умеренный некроз эпителия, инфильтраты собственной пластинки и легкая гиперплазия крипт.
Диагноз
Диарея, обезвоживание и снижение веса являются признаками кишечной инфекции. Электронная микроскопия с отрицательным контрастом выявляет вирусные частицы с пятью или шестью выступами на поверхности, напоминающими звезду. Звездообразная морфология не всегда сохраняется или четко различима.По этой причине вирусы AstV от EM можно спутать с пикорнавирусами, пикорнавирусами, энтеровирусами и энтероподобными вирусами. Во-вторых, домашние птицы могут демонстрировать клинические признаки и патологию на более поздней стадии астровирусной инфекции, но в кишечном содержимом может не быть обнаруживаемых астровирусных частиц. В такой ситуации, даже если заболевание возникло, результаты анализов могут быть отрицательными. Молекулярные методы были использованы для обнаружения и дальнейшей характеристики астровирусов. В большинстве анализов на основе ОТ-ПЦР используются праймеры, специфичные для полимеразы или гена капсида TAstV [56, 57, 96].
Смешанные вирусные инфекции
Хотя отдельные вирусы были обнаружены в стадах индейки, пораженных энтеритом, большинство исследований указывают на присутствие двух или более энтеральных вирусов [44, 45, 62, 68, 70, 75, 77, 105, 107 ]. Периодический мониторинг восьми промышленных стад индейки в США выявил постоянное присутствие TAStV и ротавируса [77]. Из 96 протестированных образцов 89,5% были положительными на астровирус и 67,7% на ротавирус. Все стада были отрицательными на TCoV, реовирус и аденовирус группы 1 во всех точках отбора проб.В другом исследовании присутствие астровирусов было зарегистрировано в 100% протестированных стад индейки [75]. TAstV-2 и TAstV-1 были обнаружены в 100 и 15,4% стад индейки соответственно. Кроме того, 12,5% стад индейки были положительными на ANV. Ротавирусы и реовирусы присутствовали в 69,7 и 45,5% стад соответственно, при этом коронавирус и аденовирус не обнаружены.
Кишечное содержимое 43 стад PES в Миннесоте было проверено на наличие энтеральных вирусов методом RT-PCR.Все 43 стада оказались положительными на ротавирус, TAstV-2 или реовирус. Во всех случаях вирусы были обнаружены либо по отдельности, либо в комбинации из 2 или 3. Восемь стай (19%) были положительными на один вирус, а остальные 35 (81%) имели комбинацию вирусов. Пятнадцать стад оказались положительными на все три вируса. Ни одно из протестированных стад не было признано положительным на TCoV [44]. Исследования, проведенные в Бразилии, показали наличие 69,7% множественных вирусов в образцах кишечника индеек. Образцы кишечника чаще всего содержали 3–4 вируса на образец, причем вирусы из семейства астровирусов (TAstV-1, TAstV-2 или ANV, или их комбинация) присутствовали во всех образцах, которые дали положительный результат на вирусы [70 ].
Возможно, что комбинация вирусов может вызвать более серьезные побочные эффекты у домашних птиц, чем отдельные вирусы. В одном исследовании индюшатина, инокулированная комбинацией TAstV-2, реовируса индейки и ротавируса, имела значительно меньшую массу тела, чем контрольная группа [91]. В экспериментальных исследованиях мы также сообщали, что пероральная инокуляция цыплят материалом PES (положительным по ротавирусу, TAstV-2 и Salmonella) приводила к значительной задержке роста на срок до 50 дней после инокуляции, и ни в какой момент после инокуляции не наблюдалось. веса оспариваемых птиц сходятся с весом контрольных.Содержимое кишечника было водянистым и пенистым / пенистым (рис.,) У цыплят, инокулированных материалом PES [49]. Точно так же индюшатины, инокулированные содержимым кишечника птиц, пораженных LTS, имели значительно меньшую массу тела в возрасте 20 недель по сравнению с индюшатинами, инокулированными содержимым кишечника птиц без LTS [68]. Такая степень замедления роста из-за энтерита домашней птицы может привести к значительным экономическим потерям для производителей индейки, поскольку пораженная птица может не достичь ожидаемого веса при продаже.На уровне фермы нельзя исключать роль агентов, отличных от этих четырех распространенных вирусов (TAstV, ротавирус, реовирус и коронавирус), в возникновении и / или повышении серьезности кишечных вирусов. Кроме того, факторы питания и менеджмент также могут играть роль в возникновении энтерита у стада. Такие шаги, как улучшение методов лечения и противомикробная терапия для минимизации потерь, имели ограниченный успех [5]. Вакцины против кишечных вирусов в настоящее время нет.
Водянистое и пенистое содержимое слепой кишки птицы, инокулированной перорально кишечным материалом птиц, пораженных синдромом энтерита домашних птиц
Водянистое и пенистое содержимое слепой кишки птицы, инокулированной перорально кишечным материалом птиц, пораженных синдромом энтерита домашних птиц.Слепая кишка разрезана
Кишечные вирусы обычно обнаруживаются с помощью ЭМ, серологии и ПЦР. Большинство из этих вирусов трудно вырастить в зародышевых яйцах курицы / индейки и клеточных культурах [95]. Таким образом, было разработано несколько анализов одиночной и множественной RT-PCR (mRT-PCR) для обнаружения астровируса индейки, ротавируса индейки, коронавируса индейки и реовируса индейки [21, 43, 56, 77, 89, 92]. Мультиплексные анализы RT-PCR экономят время и деньги, поскольку два или более вируса могут быть обнаружены в одной реакции.
Общие выводы и направления на будущее
Кишечные вирусы (TAstV, ротавирус, реовирус и коронавирус) широко распространены в стадах индейки, но их клиническое значение неясно. Полное влияние этих вирусов на производительность стада требует тщательного изучения. Хотя кишечные заболевания, вызванные вирусами, являются экономическим бременем для фермеров, выращивающих индейки, ни один вирус не появился в качестве вероятного возбудителя, чтобы можно было сформулировать соответствующие меры по профилактике и контролю [18].В нескольких исследованиях одновременно были обнаружены два или более разных вируса, что указывает на то, что несколько вирусов могут участвовать в патогенезе энтерита. Преобладание вирусов одного и того же рода в кишечнике как здоровых, так и плохо функционирующих птиц указывает на существование разных патотипов вируса. Необходимы дальнейшие исследования, чтобы определить обоснованность этих утверждений.
С появлением молекулярных диагностических тестов и секвенирования наше понимание кишечных вирусов расширилось.Также существует вероятность того, что наши текущие методы не могут обнаружить некоторые еще неизвестные вирусы / патогены, которые могут иметь последствия для энтерита индейки. Доступность и использование высокопроизводительных методов пиросеквенирования нуклеиновых кислот нового поколения, таких как секвенирование Illumina, может выявить последовательности агентов, о которых ранее не сообщалось у индеек. Метагеномные исследования РНК-вирусов в кишечнике индеек, страдающих кишечным заболеванием, выявили присутствие последовательностей вирусов дцРНК ( Reoviridae и пикобирнавирусы) и вирусов оцРНК ( Caliciviridae , Leviviridae , Picornaviridae Astroviridae и Picornaviridae ). [18].Обнаружение неизвестных вирусов методом метагеномики проложит путь к разработке методов диагностики этих вирусов. Кроме того, необходимо определить, почему одни и те же типы вируса обнаруживаются в стадах индейки, пораженных энтеритом, а также в явно здоровых стадах. Полное понимание роли кишечных вирусов и других патогенов в кишечных болезнях индеек будет иметь большое значение при разработке профилактических и контрольных мер.
Ссылки
1. Акимкин В., Биндель Ф., Хоферер М., Стинг Р., Полли Б., Ханель А., Хафез Х.М.Одношаговая RT-qPCR с системой внутреннего контроля для обнаружения ротавирусов индейки в образцах фекалий. J Virol Methods. 2011. 177 (1): 112–117. [PubMed] [Google Scholar] 2. Асано К. М., Грегори Ф., Соуза С. П., Ротава Д., Оливейра Р. Н., Вильярреал Л. Я., Рихтценхайн Л. Дж., Брандао ЧП. Ротавирус крупного рогатого скота у индеек с энтеритом. Птичий Дис. 2011; 55 (4): 697–700. [PubMed] [Google Scholar] 3. Трепет О.О., Али А., Элайш М., Ибрагим М., Мурджа М., Пантин-Джеквуд М., Саиф Ю.М., Ли К.В. Влияние коронавирусной инфекции на репродуктивную способность кур индюков.Птичий Дис. 2013; 57: 650–656. [PubMed] [Google Scholar] 4. Баньяи К., Дандар Э., Дорси К.М., Мато Т., Паля В. Геномное созвездие нового штамма ортореовируса птиц, связанного с синдромом низкорослости у бройлеров. Гены вирусов. 2011; 42: 82–89. [PubMed] [Google Scholar]5. Барнс Х. Дж., Гай Дж. С.. Синдром энтерита-смертности птицы. В: Саиф Ю.М., Барнс Х.Дж., Глиссон-младший, Фэдли А.М., Макдугалд Л.Р., Суэйн Д.Е., редакторы. Болезни птицы. 11-е изд. Эймс: Издательство государственного университета Айовы; 2003. С. 1171–1180.
6. Barnes HJ, Guy JS, Vaillancourt JP. Комплекс энтерита птицы. Rev Sci Tech. 2000. 19: 565–588. [PubMed] [Google Scholar] 7. Бенавенте Дж., Мартинес-Костас Дж. Птичий реовирус: структура и биология. Virus Res. 2007. 123: 105–119. [PubMed] [Google Scholar] 8. Безерра Д.А., да Силва Р.Р., Кайано Дж. Х., Сильвестр Р. В., де Соуза Оливейра Д., Линьярес С., Габбай Ю. Б., Маскареньяс Дж. Д.. Обнаружение ротавируса группы D птиц с использованием полимеразной цепной реакции для гена VP6. J Virol Methods. 2012; 185: 189–192. [PubMed] [Google Scholar] 9.Bidin M, Bidin Z, Majnaric D, Tisljar M, Lojkic I. Циркуляция и филогенетическая взаимосвязь астровирусов куриного и индейского происхождения, обнаруженных у домашних уток ( Anas platyrhynchos domesticus ) Avian Pathol. 2012. 41 (6): 555–562. [PubMed] [Google Scholar] 10. Боделон Дж., Лабрада Л., Мартинез-Костас Дж., Бенавенте Дж. Сегмент генома реовируса птиц S1 представляет собой функционально трицистронный ген, который экспрессирует один структурный и два неструктурных белка в инфицированных клетках. Вирусология. 2001; 290: 181–191.[PubMed] [Google Scholar] 11. Бреслин Дж. Дж., Смит Л. Г., Барнс Г. Дж., Гай Дж. С.. Сравнение процедур выделения вируса, иммуногистохимии и полимеразной цепной реакции с обратной транскриптазой для обнаружения коронавируса индейки. Птичий Дис. 2000. 44: 624–631. [PubMed] [Google Scholar] 12. Бангер А. Н., Чакон Дж. Л., Джонс Р. К., Феррейра А. Дж.. Обнаружение и молекулярная характеристика гена 3 и 5 коронавируса индейки у индюшат с тяжелым энтеритом в Бразилии. Птичий Дис. 2009. 53: 356–362. [PubMed] [Google Scholar] 13.Канелли Э., Кордиоли П., Барбьери И., Кателла А., Пеннелли Д., Церути Р., Морено А., Лавацца А. Астровирусы как возбудители энтерита домашней птицы: генетическая характеристика и лонгитюдные исследования в полевых условиях. Птичий Дис. 2012; 56: 173–182. [PubMed] [Google Scholar] 14. Cardoso TC, Castanheira TL, Teixeira MC, Rosa AC, Hirata KY, Astolphi RD, Luvizotto MC. Валидация иммуногистохимического анализа для обнаружения коронавируса индейки: быстрый и простой инструмент для скрининга в условиях ограниченных ресурсов. Poult Sci.2008. 87 (7): 1347–1352. [Бесплатная статья PMC] [PubMed] [Google Scholar] 15. Чен Ю.Н., Ву СС, Брайан Т., Хупер Т., Шредер Д., Лин Т.Л. Специфическая полимеразная цепная реакция обратной транскрипции в режиме реального времени для обнаружения и количественного определения РНК коронавируса индейки в тканях и фекалиях индеек, инфицированных коронавирусом индейки. J Virol Methods. 2010. 163: 452–458. [Бесплатная статья PMC] [PubMed] [Google Scholar] 16. Калвер Ф.А., Бриттон П., Кавана Д. Обнаружение с помощью ОТ-ПЦР птичьих коронавирусов галлиформных птиц (курица, индейка, фазан) и у попугая.Методы Мол биол. 2008; 454: 35–42. [Бесплатная статья PMC] [PubMed] [Google Scholar] 17. Да Силва SEL, Бонетти AM, Petrocelli ATM, Ferrari HF, Luvizotto MCR, Cardoso TC. Обнаружение астровируса индейки у молодняка, пораженного комплексом энтерита домашней птицы, в Бразилии. J Vet Med Sci. 2008. 70 (6): 629–631. [PubMed] [Google Scholar]19. Дэй Дж. М., Гондер Э., Дженнингс С., Ривес Д., Тилли Б., Вуминг Б. Молекулярная характеристика кишечных коронавирусов индейки, циркулирующих в США в 2012 году. Документ, представленный на ежегодном собрании Американской ассоциации патологоанатома в Чикаго, штат Иллинойс.2013.
20. Day JM, Pantin-Jackwood MJ, Spackman E. Последовательный и филогенетический анализ сегмента S1 генома реовирусов индейского происхождения. Гены вирусов. 2007. 35 (2): 235–242. [PubMed] [Google Scholar] 21. Дэй JM, Spackman E, Пантин-Джеквуд MJ. Мультиплексный ОТ-ПЦР-тест для дифференциальной идентификации астровируса индейки типа 1, астровируса индейки типа 2, астровируса цыпленка, вируса птичьего нефрита и птичьего ротавируса. Птичий Дис. 2007. 51: 681–684. [PubMed] [Google Scholar] 22. Дэй JM, Spackman E, Пантин-Джеквуд MJ.Иммунная дисфункция, вызванная реовирусом индейского происхождения, у свободных от конкретных патогенов и промышленных индюшат. Птичий Дис. 2008. 52 (3): 387–391. [PubMed] [Google Scholar] 23. Де Баттисти С., Сальвиато А., Йонассен С.М., Тоффан А., Капуа I, Каттоли Г. Генетическая характеристика астровирусов, обнаруженных у цесарок ( Numida meleagris ), выявляет отдельный генотип и предполагает межвидовую передачу между индейкой и цесаркой. Arch Virol. 2012. 157 (7): 1329–1337. [PubMed] [Google Scholar] 24. Де Бенедиктис П., Шульц-Черри С., Бернем А., Каттоли Г.Астровирусные инфекции у людей и животных — молекулярная биология, генетическое разнообразие и межвидовая передача. Заразить Genet Evol. 2011. 11 (7): 1529–1544. [Бесплатная статья PMC] [PubMed] [Google Scholar] 25. Девитт CM, Рейнольдс DL. Характеристика ротавирусов группы D. Птичий Дис. 1993; 37: 749–755. [PubMed] [Google Scholar] 26. de Wit JJ, ten Dam GB, van de Laar JMAM, Biermann Y, Verstegen I, Edens F, Schrier CC. Обнаружение и характеристика нового астровируса у кур и индеек с кишечными расстройствами и нарушениями опорно-двигательного аппарата.Avian Pathol. 2011; 40: 453–461. [PubMed] [Google Scholar] 27. Доманска-Бличарц К., Серока А., Минта З. Годовое молекулярное исследование астровирусной инфекции у индеек в Польше. Arch Virol. 2011. 156 (6): 1065–1072. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Франка М., Креспо Р., Чин Р., Вулкок П., Шивапрасад Х.Л. Ретроспективное исследование миокардита, ассоциированного с реовирусом, у индеек. Птичий Дис. 2010. 54 (3): 1026–1031. [PubMed] [Google Scholar] 30. Gomaa MH, Yoo D, Ojkic D, Barta JR. Распространенность коронавируса индейки у североамериканских индеек определяется с помощью недавно разработанного иммуноферментного анализа на основе рекомбинантного антигена.Clin Vaccine Immunol. 2008. 15 (12): 1839–1844. [Бесплатная статья PMC] [PubMed] [Google Scholar] 31. Gomaa MH, Yoo D, Ojkic D, Barta JR. Использование рекомбинантного полипептида-шипа S1 для разработки ELISA-анализа антител, специфичных к TCoV. Vet Microbiol. 2009. 138 (3–4): 281–288. [Бесплатная статья PMC] [PubMed] [Google Scholar] 32. Gomaa MH, Yoo D, Ojkic D, Barta JR. Заражение патогенным изолятом коронавируса индейки отрицательно влияет на показатели роста и морфологию кишечника молодняка индейки в Канаде. Avian Pathol.2009. 38: 279–286. [PubMed] [Google Scholar] 33. Гомес Д.Е., Хирата К.Ю., Сахеки К., Роза А.С., Лувизотто М.С., Кардосо ТК. Патология и тканевое распределение коронавируса индейки у экспериментально инфицированных цыплят и индюшат. J Comp Pathol. 2010. 143 (1): 8–13. [Бесплатная статья PMC] [PubMed] [Google Scholar] 34. Гай JS. Коронавирусный энтерит в Турции. В: Саиф Ю.М., Барнс Х.Дж., Глиссон-младший, Фэдли А.М., Макдугалд Л.Р., Суэйн Д.Е., редакторы. Болезнь птицы. 11. Эймс: Издательство государственного университета Айовы; 2003. С. 300–307. [Google Scholar] 35.Гай JS. Коронавирусный энтерит в Турции. В: Саиф YM, Fadly AM, Glisson JR, McDougald LR, Nolan LK, Swayne DE, редакторы. Болезни птицы. 12. Эймс: специалист по издательскому делу Wiley-Blackwell; 2008. С. 330–338. [Google Scholar] 36. Guy JS, Miles AM, Smith L, Fuller FJ, Schultz-Cherry S. Антигенная и геномная характеристика энтеровирусоподобного вируса индейки (Северная Каролина, изолят 1988 г.): идентификация вируса как астровируса индейки 2. Avian Dis. 2004. 48 (1): 206–211. [PubMed] [Google Scholar] 37.Гай Дж. С., Смит Л. Г., Бреслин Дж. Дж., Пакпиньо С. Разработка конкурентного иммуноферментного анализа для обнаружения антител к коронавирусу индейки. Птичий Дис. 2002. 46 (2): 334–341. [PubMed] [Google Scholar] 38. Гай Дж. С., Смит Л. Г., Бреслин Дж. Дж., Вайланкур Дж. П., Барнс Х. Дж. Высокая смертность и подавление роста, экспериментально вызванные у молодых индеек двойным заражением энтеропатогенным вирусом Escherichia coli и коронавирусом индейки. Птичий Дис. 2000. 44: 105–113. [PubMed] [Google Scholar] 39.Heggen-Peay CL, Qureshi MA, Edens FW, Sherry B, Wakenell PS, O’Connell PH, Schat KA. Выделение реовируса от синдрома энтерита и смертности домашних птиц и его патогенность у индюшат. Птичий Дис. 2002. 46 (1): 32–47. [PubMed] [Google Scholar] 40. Имада Т., Ямагути С., Масе М, Цукамото К., Кубо М., Морока А. Вирус птичьего нефрита (ANV) как новый член семейства Astroviridae и конструирование инфекционной кДНК ANV. J Virol. 2000; 74: 8487–8493. [Бесплатная статья PMC] [PubMed] [Google Scholar] 41.Исмаил М.М., Тан А.Ю., Саиф Ю.М. Патогенность коронавируса индейки у индеек и кур. Птичий Дис. 2003. 47: 515–522. [PubMed] [Google Scholar] 42. Джеквуд М.У., Бойнтон Т.О., Хилт Д.А., Маккинли Е.Т., Киссинджер Дж.С., Патерсон А.Х., Робертсон Дж., Лемке С., МакКолл А.В., Уильямс С.М., Джеквуд Дж. В., Берд Л.А. Возникновение коронавируса группы 3 в результате рекомбинации. Вирусология. 2010; 398: 98–108. [Бесплатная статья PMC] [PubMed] [Google Scholar] 43. Джиндал Н., Чандер И., Патнаяк Д.П., Мор СК, Зиглер А.Ф., Гоял С.М. Мультиплексная RT-PCR для обнаружения астровирусов, ротавирусов и реовирусов у индеек.Птичий Дис. 2012; 56: 592–596. [PubMed] [Google Scholar] 44. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М. Выявление и молекулярная характеристика кишечных вирусов синдрома энтерита домашней птицы у индеек. Poult Sci. 2010. 89: 217–226. [Бесплатная статья PMC] [PubMed] [Google Scholar] 45. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М. Обнаружение и молекулярная характеристика кишечных вирусов у племенных индюков. Avian Pathol. 2010; 39: 53–61. [PubMed] [Google Scholar] 46. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М.Сравнение последовательностей капсидных генов астровируса-2 индейки у индюшат, пораженных синдромом энтерита, и у практически здоровых индюшат. Arch Virol. 2011; 156: 969–977. [PubMed] [Google Scholar] 47. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Ретроспективное исследование синдрома энтерита домашней птицы в Миннесоте. Птичий Дис. 2009; 53: 268–275. [PubMed] [Google Scholar] 48. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Экспериментальное воспроизведение синдрома энтерита домашней птицы: клинические данные, реакция роста и микробиология.Poult Sci. 2009; 88: 949–958. [Бесплатная статья PMC] [PubMed] [Google Scholar] 49. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Продолжительность угнетения роста и выделения патогенов при синдроме энтерита экспериментально воспроизведенных индюшат. Птичий Дис. 2009; 53: 517–522. [PubMed] [Google Scholar] 50. Johne R, Otto P, Roth B, Löhren U, Belnap D, Reetz J, Trojnar E. Анализ последовательности сегмента генома, кодирующего VP6, ротавирусов птичьей группы F и G. Вирусология. 2011; 412: 384–391. [PubMed] [Google Scholar] 51.Джотикумар Н, Канг Г., Хилл ВР. Широкореактивный анализ TaqMan для обнаружения ротавируса в режиме реального времени с помощью ОТ-ПЦР в клинических образцах и образцах окружающей среды. J Virol Methods. 2009. 155: 126–131. [PubMed] [Google Scholar] 52. Капчинский Д.Р., Селлерс Х.С., Симмонс В., Шульц-Черри С. Анализ последовательности гена S3 из реовируса индейки. Гены вирусов. 2002. 25 (1): 95–100. [Бесплатная статья PMC] [PubMed] [Google Scholar] 53. Кэ GM, Cheng HL, Ke LY, Ji WT, Chulu JL, Liao MH, Chang TJ, Liu HJ. Разработка количественного светового циклера RT-PCR в реальном времени для обнаружения птичьего реовируса.J Virol Methods. 2006; 133 (1): 6–13. [PubMed] [Google Scholar] 54. King AMQ, Lefkowiz E, Adams M, Carstens E. Таксономия вирусов: девятый доклад международного комитета по таксономии вирусов. Уолтем: Academic Press; 2012. [Google Scholar] 55. Koci MD, Schultz-Cherry S. Птичьи астровирусы. Avian Pathol. 2002; 31: 213–227. [PubMed] [Google Scholar] 56. Koci MD, Seal BS, Schultz-Cherry S. Разработка диагностического теста RT-PCR для птичьего астровируса. J Virol Methods. 2000. 90: 79–83. [Бесплатная статья PMC] [PubMed] [Google Scholar] 58.Кул Д.А., Холмс И.Х. Нуклеотидная и расчетная аминокислотные последовательности Ty-1 Vp7 ротавируса птиц значительно отличаются от таковых ротавируса Ch-2. Arch Virol. 1993. 129 (1–4): 227–234. [PubMed] [Google Scholar] 59. Lin TL, Loa CC, Tsai SC, Wu CC, Bryan TA, Thacker HL, Hooper T., Schrader D. Характеристика коронавируса индейки у индюшат с острым энтеритом. Vet Microbiol. 2002. 84 (1–2): 179–186. [Бесплатная статья PMC] [PubMed] [Google Scholar] 60. Линь Т.Л., Лоа СС, Ву СС. Полные последовательности 3′-концевой кодирующей области для генов структурных белков коронавируса индейки.Virus Res. 2004. 106 (1): 61–70. [Бесплатная статья PMC] [PubMed] [Google Scholar] 61. Loa CC, Wu CC, Lin TL. Сравнение 3′-концевых кодирующих областей изолятов коронавируса индейки из Индианы, Северной Каролины и Миннесоты со штаммами коронавируса инфекционного бронхита кур. Интервирология. 2006. 49 (4): 230–238. [Бесплатная статья PMC] [PubMed] [Google Scholar] 62. Lojkic I, Bidin M, Bidin Z, Mikec M. Вирусные агенты, связанные с энтеритом домашней птицы в хорватских промышленных стадах индейки. Acta Vet BRNO. 2010; 79: 91–98.[Google Scholar] 63. Маттейнссенс Дж., Мартелла В., Ван Ранст М. Оценка приоритетной статьи: геномная эволюция, межвидовой барьер хозяина, реассортировка и классификация ротавирусов. Будущий Virol. 2010. 5: 385–390. [Google Scholar] 64. Matthijnssens J, Otto PH, Ciarlet M, Desselberger U, Van Ranst M, Johne R. Значения отсечения на основе последовательности VP6 в качестве критерия для демаркации видов ротавирусов. Arch Virol. 2012. 157 (6): 1177–1182. [PubMed] [Google Scholar] 65. Maurel S, Toquin D, Briand FX, Queguiner M, Allee C, Bertin J, Ravillion L, Retaux C, Turblin V, Morvan H, Eterradossi N.Первые полноразмерные последовательности гена S европейских изолятов выявили дальнейшее разнообразие коронавирусов индейки. Avian Pathol. 2011; 40: 179–189. [PubMed] [Google Scholar] 66. Макналти М.С., Аллан Г.М., Маккракен Р.М. Экспериментальное заражение цыплят ротавирусами: клинические и вирусологические данные. Avian Pathol. 1983; 12: 45–54. [PubMed] [Google Scholar] 67. Мор С.К., Абин М., Коста Дж., Дуррани А., Джиндал Н., Гоял С.М., Патнаяк Д.П. Роль астровируса индейки 2 типа в синдроме энтерита домашних птиц. Poult Sci.2011; 90: 2747–2752. [Бесплатная статья PMC] [PubMed] [Google Scholar] 68. Мор С.К., Шарафельдин Т.А., Абин М., Кромм М., Портер Р.Э., Гоял С.М., Патнаяк Д.П. Возникновение кишечных вирусов при синдроме легкой индейки. Avian Pathol. 2013. 42 (5): 497–501. [PubMed] [Google Scholar] 69. Мор С.К., Шарафельдин Т.А., Портер Р.Э., Зиглер А., Патнаяк Д.П., Гоял С.М. Выделение и характеристика реовируса артрита индейки. Птичий Дис. 2013. 57 (1): 97–103. [PubMed] [Google Scholar] 70. Моура-Альварес Дж., Чакон СП, Сканавини Л.С., Нуньес ЛФН, Астольфи-Феррейра К.С., Джонс Р.К., Пиантино Феррейра А.Дж.Кишечные вирусы в стадах бразильских индейок: частота единичных и множественных вирусных инфекций в зависимости от возраста и клинических признаков кишечного заболевания. Poult Sci. 2013; 92: 945–955. [Бесплатная статья PMC] [PubMed] [Google Scholar] 71. Otto PH, Ahmed MU, Hotzel H, Machnowska P, Reetz J, Roth B, Trojnar E, Johne R. Обнаружение птичьих ротавирусов групп A, D, F и G у больных кур и индеек из Европы и Бангладеш. Vet Microbiol. 2012; 156 (1-2): 8-15. [Бесплатная статья PMC] [PubMed] [Google Scholar] 72.Отто П., Либлер-Тенорио Е.М., Эльшнер М., Ритц Дж., Лорен У., Диллер Р. Обнаружение ротавирусов и поражений кишечника у цыплят-бройлеров в стадах с синдромом короткого роста и задержки роста (RSS) Avian Dis. 2006; 50: 411–418. [PubMed] [Google Scholar] 73. Пакпиньо С., Лей Д.Х., Барнс Х.Дж., Вайланкур Дж. П., Гай Дж. С.. Распространенность энтеропатогенной Escherichia coli в естественных случаях синдрома энтерита-смертности домашних птиц. Птичий Дис. 2002. 46 (2): 360–369. [PubMed] [Google Scholar] 74. Palade EA, Demeter Z, Hornyak A, Nemes C, Kisary J, Rusvai M.Высокая распространенность парвовируса индейки в стадах индейки из Венгрии, страдающих синдромами кишечных заболеваний. Птичий Дис. 2011; 55 (3): 468–475. [PubMed] [Google Scholar] 75. Пантин-Джеквуд MJ, Day JM, Jackwood MW, Spackman E. Кишечные вирусы, обнаруженные молекулярными методами в коммерческих стадах кур и индейок в Соединенных Штатах в период с 2005 по 2006 гг. Птичий Дис. 2008. 52: 235–244. [PubMed] [Google Scholar] 76. Пантин-Джеквуд MJ, Spackman E, Day JM. Патогенез астровирусов индейки 2 типа с вариантными генами капсида у 2-дневных цыплят, свободных от специфических патогенов.Avian Pathol. 2008; 37: 193–201. [PubMed] [Google Scholar] 77. Пантин-Джеквуд MJ, Spackman E, Day JM, Rives D. Периодический мониторинг промышленных индеек на предмет энтеросолюбии указывает на постоянное присутствие астровируса и ротавируса на фермах. Птичий Дис. 2007. 51: 674–680. [PubMed] [Google Scholar] 78. Пантин-Джеквуд MJ, Spackman E, Woolcock PR. Молекулярная характеристика и типирование астровирусов кур и индейки, циркулирующих в США: значение для диагностики. Птичий Дис. 2006; 50: 397–404.[PubMed] [Google Scholar] 79. Пантин-Джеквуд MJ, Strother KO, Mundt E, Zsak L, Day JM, Spackman E. Молекулярная характеристика птичьих астровирусов. Arch Virol. 2011; 156: 235–244. [PubMed] [Google Scholar] 80. Куреши М.А., Саиф Ю.М., Хегген-Пи К.Л., Эденс Ф.В., Хавенштейн ГБ. Индукция функциональных дефектов в макрофагах астровирусом индейки, ассоциированным с энтеритом и синдромом смертности домашних птиц. Птичий Дис. 2001. 45 (4): 853–861. [PubMed] [Google Scholar] 81. Рамиг РФ, Ciarlet M, Mertens PPC, Dermody TS. Ротавирус.В: Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA, редакторы. Таксономия вирусов: восьмой отчет международного комитета по таксономии вирусов. Амстердам: Academic Press; 2005. С. 484–496. [Google Scholar] 82. Рейнольдс Д.Л., Саиф Ю.М. Астровирус: причина кишечного заболевания у индюшат. Птичий Дис. 1986; 30: 728–735. [PubMed] [Google Scholar]83. Роббинс К.М. Эпидемиология и диагностика коронавируса в Турции. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс.2013.
84. Родригес-Диас Дж., Рубилар-Абреу Э., Шпицнер М., Хедлунд К.О., Липранди Ф., Свенссон Л. Дизайн мультиплексной вложенной ПЦР для генотипирования NSP4 ротавируса группы А. J Virol Methods. 2008. 149: 240–245. [PubMed] [Google Scholar] 85. Ruff MD, Rosenberger JK. Сопутствующие инфекции реовирусами и кокцидиями у бройлеров. Птичий Дис. 1985. 29 (2): 465–478. [PubMed] [Google Scholar] 86. Шульц-Черри С., Капчинский Д.Р., Симмонс В.М., Кочи М.Д., Браун С., Барнс Х.Дж. Идентификация агента (ов), связанного с синдромом смертности от энтерита домашней птицы: важность тимуса.Птичий Дис. 2000. 44: 256–265. [PubMed] [Google Scholar] 87. Шуман Т., Хотцель Х., Отто П., Джон Р. Доказательства межвидовой передачи и реассортации среди ротавирусов птичьей группы А. Вирусология. 2009. 386 (2): 334–343. [PubMed] [Google Scholar] 88. Продавцы HS, Linnemann EG, Pereira L, Kapczynski DR. Филогенетический анализ гена белка сигма 2 реовирусов индейки. Птичий Дис. 2004. 48 (3): 651–657. [PubMed] [Google Scholar] 89. Продавцы HS, Koci MD, Linnemann E, Kelley LA, Schultz-Cherry S. Разработка диагностического теста мультиплексной полимеразной цепной реакции с обратной транскрипцией, специфичного для астровируса и коронавируса индейки.Птичий Дис. 2004. 48: 531–539. [PubMed] [Google Scholar] 90. Shivaprasad HL, Franca M, Woolcock PR, Nordhausen R, Day JM, Pantin-Jackwood M. Миокардит, связанный с реовирусом у индюшат. Птичий Дис. 2009. 53: 523–532. [PubMed] [Google Scholar] 91. Spackman E, Day JM, Pantin-Jackwood MJ. Сопутствующие инфекции астровируса, реовируса и ротавируса вызывают снижение прибавки в весе у широкогрудых белых цыплят. Птичий Дис. 2010; 54: 16–21. [PubMed] [Google Scholar] 92. Спакман Э., Капчинский Д., Селлерс Х. Мультиплексная полимеразная цепная реакция обратной транскрипции в реальном времени для обнаружения трех вирусов, связанных с комплексом энтерита домашней птицы: астровируса индейки, коронавируса индейки и реовируса индейки.Птичий Дис. 2005; 49: 86–91. [PubMed] [Google Scholar] 93. Спакман Э., Пантин-Джеквуд М., Дэй Дж. М., Селлерс Х. Патогенез реовирусов индейского происхождения у индеек и кур. Avian Pathol. 2005; 34: 291–296. [PubMed] [Google Scholar] 94. Штамм E, Kelley LA, Schultz-Cherry S, Muse SV, Koci MD. Геномный анализ близкородственных астровирусов. J Virol. 2008. 82: 5099–5103. [Бесплатная статья PMC] [PubMed] [Google Scholar] 95. Сугияма М.К., Гото К., Уэмукай Х., Мори Й., Ито Н., Минамото Н. Присоединение и инфицирование к клеткам MA104 ротавирусов птиц требует присутствия сиаловой кислоты на поверхности клетки.J Vet Med Sci. 2004. 66: 461–463. [PubMed] [Google Scholar] 96. Тан Й., Исмаил М.М., Саиф Ю.М. Разработка иммуноферментного анализа с захватом антигена и ОТ-ПЦР для обнаружения астровирусов индейки. Птичий Дис. 2005. 49 (2): 182–188. [PubMed] [Google Scholar] 97. Тан Й., Мурджа А.М., Саиф Ю.М. Молекулярная характеристика гена капсида двух серотипов астровирусов индейки. Птичий Дис. 2005. 49 (4): 514–519. [PubMed] [Google Scholar] 98. MC Teixeira, MC Luvizotto, Ferrari HF, Mendes AR, da Silva SE, Cardoso TC.Обнаружение коронавируса индейки у коммерческих индюшат в Бразилии. Avian Pathol. 2007. 36 (1): 29–33. [PubMed] [Google Scholar] 99. Тейл К.В., Саиф Ю.М. Возрастные инфекции ротавируса, ротавирусоподобного вируса и атипичного ротавируса в стадах индейки. J Clin Microbiol. 1987. 25 (2): 333–337. [Бесплатная статья PMC] [PubMed] [Google Scholar]100. Тилли Б.Дж., Гондер Э., Мейсон С., Уикс Дж. Вспышка коронавируса в Турции в Северной Каролине. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс.2013.
101. Тоффан А., Катания С., Сальвиато А., Де Баттисти С., Васкеллари М., Тосон М., Капуа I, Каттоли Г. Экспериментальное заражение домашних птиц и цесарок генетически отличными птичьими астровирусами. Avian Pathol. 2012; 41: 429–435. [PubMed] [Google Scholar] 102. Trojnar E, Otto P, Roth B, Reetz J, Johne R. Сегменты генома ротавируса группы D имеют консервативные концы группы A, но кодируют специфичные для группы белки. J Virol. 2010; 84: 10254–10265. [Бесплатная статья PMC] [PubMed] [Google Scholar] 103.Trojnar E, Sachsenroder J, Twardziok S, Reetz J, Otto PH, Johne R. Идентификация ротавируса птичьей группы А, содержащего новый ген VP4, близкий родству к генам ротавирусов млекопитающих. J Gen Virol. 2013; 94: 136–142. [PubMed] [Google Scholar] 104. Велаюдхан Б.Т., Шин Х.Д., Лопес В.К., Хупер Т., Халворсон Д.А., Нагараджа К.В. Анализ обратной транскриптазы-полимеразной цепной реакции для диагностики коронавирусной инфекции индейки. J Vet Diagn Invest. 2003. 15 (6): 592–596. [PubMed] [Google Scholar] 105.Woolcock PR, Shivaprasad HL. Электронно-микроскопическая идентификация вирусов, связанных с энтеритом домашней птицы, у индеек, выращенных в Калифорнии в 1993–2003 гг. Птичий Дис. 2008. 52: 209–213. [PubMed] [Google Scholar]106. Вуминг Б. Вспышка коронавируса на северо-западе Арканзаса. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс. 2013.
107. Ю М., Исмаил М. М., Куреши М. А., Дерт Р. Н., Барнс Х. Дж., Саиф Ю. М.. Вирусные агенты, связанные с энтеритом и синдромом смертности домашних птиц: роль небольшого круглого вируса и коронавируса индейки.Птичий Дис. 2000. 44 (2): 297–304. [PubMed] [Google Scholar] 108. Ю М, Тан Й, Го М, Чжан Кью, Саиф Ю. Характеристика небольшого круглого вируса, связанного с синдромом энтерита и смертности цыплят. Птичий Дис. 2000. 44 (3): 600–610. [PubMed] [Google Scholar] 109. Zsak L, Strother KO, Kisary J. Анализ частичной последовательности генома парвовирусов, связанных с кишечным заболеванием у домашней птицы. Avian Pathol. 2008. 37 (4): 435–441. [PubMed] [Google Scholar]Кишечных вирусов энтерита индейки
Вирусная болезнь.2014 июн; 25 (2): 173–185.
, , иНареш Джиндал
Департамент ветеринарного здравоохранения и эпидемиологии, Колледж ветеринарных наук, Университет ветеринарии и зоотехники Лала Ладжпат Рай, Хисар, 125 004 Индия
9004, Индия. К. МорКафедра ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Сент-Пол, Миннесота 55108 США
Сагар М.Гоял
Департамент ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Сент-Пол, Миннесота 55108 США
Департамент ветеринарного здравоохранения и эпидемиологии, Колледж ветеринарных наук, Ветеринарный университет Лала Ладжпат Рай. Науки о животных, Хисар, 125 004 Индия
Департамент ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Сент-Пол, Миннесота 55108 США
Автор, отвечающий за переписку.Поступило 2 октября 2013 г .; Принято 27 января 2014 г.
Эта статья цитируется в других статьях в PMC.Abstract
Здоровье кишечника очень важно для получения максимальной отдачи с точки зрения увеличения веса и яйценоскости. Кишечные заболевания, такие как комплекс энтерита домашней птицы (PEC) у индеек, не позволяют максимально реализовать их производственный потенциал. Был вовлечен ряд вирусов, бактерий и простейших, но первичная этиология окончательно не установлена. Раньше электронная микроскопия использовалась для обнаружения кишечных вирусов, которые определялись исключительно на основе их морфологии.С появлением методов быстрой молекулярной диагностики и секвенирования нуклеиновых кислот нового поколения исследователи добились больших успехов в идентификации и характеристике вирусов, связанных с PEC. Молекулярные методы также помогли нам идентифицировать патогены, которые ранее не были известны. Региональные и национальные исследования выявили наличие нескольких различных кишечных вирусов в PEC, включая ротавирус, астровирус, реовирус и коронавирус, по отдельности или в комбинации.По-прежнему могут существовать неизвестные патогены, которые могут прямо или косвенно играть роль в энтерите у индеек. Этот обзор будет посвящен роли коронавируса, ротавируса, реовируса и астровируса индейки в энтерите индейки.
Ключевые слова: Энтерит, Индейки, Коронавирус, Ротавирус, Реовирус, Астровирус
Введение
На протяжении многих лет у индеек было описано несколько кишечных заболеваний, а именно: коронавирусный энтерит индеек, синдром мальдигестии, синдром ранения и задержки роста у индеек, синдром мальабсорбции у домашней птицы, синдром энтерита и смертности домашних птиц (PEMS), пиковая смертность индеек и вирусный энтерит индейки.Все эти синдромы были включены в комплекс энтерита домашней птицы (PEC), общий термин, обозначающий инфекционные кишечные заболевания молодняка индеек [6]. ПЭК — это экономически важное заболевание молодняка индеек, характеризующееся энтеритом, диареей, умеренной или выраженной депрессией роста, нарушением использования корма, плохой прибавкой веса и в некоторых случаях высокой смертностью, как при ПЕМС [6]. Ряд вирусов (коронавирус, калицивирус, реовирус, астровирус, ротавирус, пикорнавирус, пикобирнавирус, парвовирус и аденовирус), бактерий ( Escherichia coli , Salmonella spp., Clostridia, Campylobacter и Enterococcus ) и простейшие (кокцидии и криптоспоридиумы) участвуют в PEC [6, 18, 47, 73, 105, 109]. Недавно у индюшат Миннесоты были описаны два новых синдрома, а именно «синдром энтерита домашней птицы», или PES, и «синдром легкой индейки», или LTS. ПЭС — это инфекционное кишечное заболевание молодых индеек в возрасте от 1 дня до 7 недель, которое характеризуется диареей, депрессией и летаргией с бледным кишечником и / или чрезмерно жидким содержимым слепой кишки [47].LTS — это проблема индеек товарного возраста, имеющих меньшую массу тела по сравнению с их стандартными породными характеристиками [68]. Мы полагаем, что энтерит у домашних птиц в молодом возрасте создает условия для будущего развития LTS у более старых птиц. Кроме того, мы рассматриваем как PES, так и LTS как часть PEC.
Кишечные вирусы являются частой причиной первичного повреждения желудочно-кишечного тракта (ЖКТ) молодых цыплят, создавая благоприятную среду для роста бактерий и / или простейших и вызывая дальнейшее повреждение кишечника.Возможно, что повреждение кишечника сначала кишечными вирусами, а затем вторичными патогенами может привести к необратимым изменениям в организме хозяина, что может привести к снижению массы тела при продаже. Цель этого обзора — подробно обсудить четыре наиболее важных вируса, вызывающих энтерит индейки, а именно коронавирус, реовирус, ротавирус и астровирус.
Коронавирус
Коронавирусный энтерит индейки — острое заразное заболевание, характеризующееся тупостью, депрессией, диареей и сниженным набором массы тела.Заболевание было впервые описано в 1940-х годах в штате Вашингтон, США. Возбудитель не был идентифицирован, и заболевание было названо «грязевой лихорадкой». Однако после выявления в этих случаях коронавируса индейки (TCoV) название было изменено на «коронавирусный энтерит индеек». В 1970-х годах эта болезнь имела самые разрушительные последствия для индеек из Миннесоты. Были предприняты шаги по искоренению болезни, которые привели к элиминации этого вируса из Миннесоты. В настоящее время TCoV спорадически регистрируется из регионов Северной Америки и других стран, где выращивают индейку.TCoV также связан с PEMS, острым высококонтагиозным кишечным заболеванием, характеризующимся депрессией, анорексией, диареей и высокой смертностью индеек.
Геном
Коронавирусы представляют собой плеоморфные частицы с оболочкой, примерно сферической формы, диаметром от 50 до 200 нм. Согласно последнему предложению Международного комитета по таксономии вирусов (ICTV), семейство Coronaviridae включает два подсемейства: Coronavirinae и Torovirinae. Coronavirinae состоит из трех родов: Alphacoronavirus , Betacoronavirus и Gammacoronavirus [54], из которых последний включает коронавирусы птиц. Геном коронавируса состоит из одноцепочечной РНК с положительным смыслом. Полноразмерный геном состоит из 27 632 нуклеотидов плюс 3 ‘поли (А) хвост. Есть две открытые рамки считывания (ORF), ORF 1a и 1b, которые находятся в первых двух третях генома с девятью дополнительными ORF ниже по течению.Область между генами белков мембраны (M) и нуклеокапсида (N) содержит три потенциальных малых ORF: (1) ORF-X, (2) ранее не охарактеризованную ORF со связанной предполагаемой TRS в гене M (очевидно, общей для всей группы III coronaviruses) и (3) ранее описанные ORF 5a и 5b. TCoV не содержит гена гемагглютинин-эстеразы [29].
Сообщалось о вариабельности в различных генных сегментах TCoV. Сравнение последовательностей полной 3′-концевой области, кодирующей структурный белок четырех TCoV, с соответствующими последовательностями вируса инфекционного бронхита (IBV) показало, что TCoV, вероятно, имеет то же происхождение, что и IBV, и приобретенные последовательности S-гена для тропизма кишечника индейки во время процесс эволюции [61].Lin et al. [60] сообщили, что последовательность S-белка TCoV имеет только 33,8–33,9% идентичности с IBV, хотя оба вируса сгруппированы в одну и ту же геномную линию. Полноразмерные последовательности гена spike (S) французского происхождения TCoV обладали 98% идентичностью как на уровне нуклеотидов, так и на уровне аминокислот [65]. Однако эти вирусы показали 65 и 60% сходства с североамериканскими TCoV и 50 и 37% сходства с IBV на уровне нуклеотидов и аминокислот, соответственно.
Заболеваемость и распространение
В отличие от обычно неопределенных синдромов вирусных кишечных заболеваний, TCoV-энтерит можно напрямую отнести к инфекции, вызванной TCoV [35, 38, 41].TCoV распространен во всем мире и был обнаружен у пораженных энтеритом индеек из Северной Америки, Южной Америки и Европы [34, 59, 65, 70, 98]. Было обнаружено, что 37% кишечных проб от больных стад индейки в Европе оказались положительными на TCoV [65]. В недавнем исследовании, проведенном в Бразилии, большинство образцов кишечника, взятых как у пораженных энтеритом, так и у здоровых индеек, оказались положительными на TCoV [70]. В этом исследовании 55,3% TCoV-положительных образцов также были положительными на другие кишечные вирусы.Недавно были зарегистрированы вспышки коронавирусного энтерита из Арканзаса и Северной Каролины в США с тяжелой диареей и смертностью [19, 83, 100, 106], что указывает на важность постоянного эпидемиологического надзора за этим вирусом. TCoV, по-видимому, отсутствует в синдромах ПЭК без ПЭМС [44, 45, 62, 68, 75, 105]. Например, коронавирус не был обнаружен, когда в Калифорнии были протестированы 2400 образцов от домашних птиц с клиническими признаками энтерита [105]. Аналогичным образом, в исследовании 33 стай индеек, пораженных энтеритом, в США, Pantin-Jackwood et al.[75] не нашли TCoV. Мы также не обнаружили TCoV в случаях PES и LTS [44, 45, 68].
TCoV может поражать птиц любой возрастной группы, но вызывает высокую смертность молодых индюшат в возрасте 1–4 недель. Смертность колеблется от менее 10 до 50%. Индюки, по-видимому, являются единственным естественным хозяином TCoV; при экспериментальной инокуляции TCoV вызывал заболевание у домашних птиц, но не у цыплят [42]. Репликация TCoV происходит в энтероцитах, выстилающих апикальную часть ворсинок кишечника. Разрушение эпителия ворсинок кишечника приводит к диарее, нарушению пищеварения и мальабсорбции, что может привести к снижению прибавки в весе у индюшат.
Передача
Вирус присутствует в основном в кишечнике и фекалиях пораженных домашних птиц, следовательно, передача вируса осуществляется фекальным оральным путем. Прямая передача происходит при прямом контакте с инфицированными птицами, тогда как косвенная передача происходит механическим путем через рабочих и / или зараженное оборудование. Однако в экспериментальном исследовании антиген TCoV также может быть обнаружен в придаточных пазухах носа и слезной добавочной железе (железе Хардера) инфицированных цыплят в течение 14 дней после инокуляции [33].Вирус выделяется с фекалиями индеек в течение многих недель после выздоровления от клинического заболевания. Большинство случаев заражения происходит летом (май – август), иногда осенью. TCoV может сохраняться в течение длительного периода времени и может легко передаваться между птицефабриками в прохладной окружающей среде.
Клинические признаки и патология
Индюки, пораженные TCoV, сбиваются в кучу и демонстрируют взъерошенные перья, снижение потребления корма и воды, влажный помет и потерю живой массы.У пожилых птиц может наблюдаться депрессия, диарея и задержка роста [34]. Помет водянистый и пенистый, может содержать слизь, от зеленого до коричневого цвета. Клинические признаки могут сохраняться до 2 недель, а восстановление веса, если оно будет достигнуто, может занять несколько недель. У несушек наблюдается снижение яйценоскости [3] из-за аномальной пигментации яиц. Птицы в возрасте от 1 до 4 недель, у которых наблюдаются диарея, обезвоживание и задержка роста на 40% или более, считаются страдающими PEMS. Существует две формы PEMS: (1) тяжелая форма i.е. рост нравственности индеек (SMT), который определяется как смертность, равная или превышающая 1% в течение 3 дней подряд или 9% в возрасте от 8 до 28 дней; и (2) легкая форма, которая определяется как смертность более 2%, но менее 9% в течение того же периода времени, но не равная или не превышающая 1% в течение трех дней подряд. В экспериментальном исследовании, как заболеваемость, так и смертность регистрировались у домашних птиц, экспериментально инокулированных TCoV в возрасте 2 дней, однако птицы, инфицированные в возрасте 28 дней, имели значительную задержку роста, но не демонстрировали смертности [32].
TCoV может действовать синергетически с другими патогенами, увеличивая тяжесть заболевания, или может предрасполагать птиц к вторичным инфекциям. У цыплят, инокулированных только TCoV, развилась умеренная депрессия роста без гибели, а у свиней, инокулированных только энтеропатогенной Escherichia coli (EPEC) , клиническое заболевание не развилось. Однако инокуляция цыплят TCoV и EPEC одновременно вызвала повышение смертности, подавление роста и прикрепление / стирание повреждений [38].
У пораженных птиц наблюдается задержка роста и обезвоживание. Кишечник заметно увеличен и заполнен рыхлым желтым содержимым. Стенки кишечника вялые и бледные. Caeca также вздуты с пенистым и водянистым содержимым. Крупные изменения, такие как атрофия поджелудочной железы, селезенки и бурсы Фабрициуса, также могут наблюдаться наряду с расширением желчного пузыря и мочеточников. У инфицированных кур наблюдаются более выраженные поражения яичников с деформированными, плохо развитыми яичниками и скоплением казеозного экссудата в брюшине.Также наблюдаются спайки брюшины яйцевода.
В зависимости от тяжести инфекции наблюдается от легкого мультифокального до тяжелого энтерита с атрофией ворсинок. Наблюдается слущивание эпителиальных клеток с застоем и обширной инфильтрацией лимфоцитов и гетерофилов в собственную пластинку ворсинок. Кроме того, также может наблюдаться истощение фолликулярных лимфоидов и атрофия коркового вещества тимуса.
Диагноз
Клинические признаки, такие как диарея, депрессия, скученность и задержка роста, указывают на кишечную инфекцию.Высокая смертность молодняка может указывать на ПЕМС.
Выделение вируса, электронная микроскопия (ЭМ), обнаружение вирусной РНК и серология могут помочь в диагностике. Вирус можно выращивать в зародышах индейки или куриных яйцах, но его трудно выращивать в клеточных культурах. ЭМ с отрицательным контрастом может выявить вирусные частицы размером ~ 50–200 нм с типичной морфологией в форме короны в содержимом кишечника. Прямые и непрямые флуоресцентные тесты на антитела и ELISA использовались для обнаружения антигена TCoV и антител соответственно [30, 31, 37].О высокой распространенности TCoV сообщалось от заводчиков (71,1%) и мясных индеек (56,7%) в Онтарио, Канада [31]. В Арканзасе 64,2% образцов сыворотки дали положительный результат на антитела [30].
Доступны молекулярные тесты для обнаружения и дифференциации TCoV от других кишечных вирусов. Доступны различные тесты полимеразной цепной реакции с обратной транскрипцией (RT-PCR), нацеленные на 3′-нетранслируемую область (UTR), S, N и / или M ген TCoV [11, 12, 16, 89, 92, 104]. Также была разработана ОТ-ПЦР в реальном времени, которая является наиболее чувствительным и специфическим тестом, с помощью которого можно количественно определить вирусную нагрузку в тканях индейки, а также в образцах фекалий [15].Прямая иммуногистохимия была рекомендована в качестве альтернативы ОТ-ПЦР для обнаружения TCoV в полевых образцах [14].
Профилактика и контроль
Специального лечения или вакцины нет. Биобезопасность, при которой особое внимание уделяется управлению подстилкой и надлежащей утилизации мертвых птиц и использованного подстилки, похоже, единственный способ предотвратить распространение инфекции. Кроме того, общие меры, такие как уменьшение влажности подстилки, использование антибиотиков для борьбы с вторичными бактериальными инфекциями и улучшение гигиены, могут минимизировать последствия энтерита TCoV.
Реовирус
Птичий реовирус был впервые выделен от кур, страдающих хроническим респираторным заболеванием, и получил название возбудитель Фейи-Кроули (FC). Позже возбудитель FC был охарактеризован как реовирус. Однако у кур птичий реовирус вызывает множество синдромов, включая вирусный артрит / теносиновит. Недавно реовирусы были выделены от артрита / тендосиновита индейки и получили название TARV (реовирус артрита индейки) [69]. Реовирусы в основном бывают двух типов: фузогенные и нефузогенные.Слитые вирусы обладают способностью вызывать слияние инфицированных клеток, что приводит к образованию многоядерных синцитий, в то время как нефузогенные вирусы не вызывают слияния инфицированных клеток. К нефузогенным типам относятся, в основном, реовирусы млекопитающих (MRV), тогда как слитные реовирусы поражают млекопитающих, птиц и рептилий.
Геном
Реовирус принадлежит к роду Orthoreovirus в семействе Orthoreoviridae. Вирус без оболочки, имеет икосаэдрическую симметрию и размер частиц 70–80 нм.Он содержит геном двухцепочечной (ds) РНК из десяти сегментов, которые делятся на три класса, называемые большими (L), средними (M) и малыми (S), в зависимости от их характера миграции при электрофорезе в полиакриламидном геле. Гены L и M далее подразделяются на три сегмента каждый (L1, L2, L3 и M1, M2, M3, соответственно), в то время как ген S имеет четыре сегмента (S1, S2, S3, S4) [7].
Геном реовируса имеет 12 открытых рамок считывания, которые кодируют восемь структурных и четыре неструктурных белка.Структурные белки являются важной частью вирионов потомства, в то время как неструктурные белки не присутствуют в зрелом вирионе и экспрессируются только в инфицированных клетках. Структурными белками, кодируемыми генами L, M и S, являются лямбда (λ), мю (μ) и сигма (σ) соответственно. Три структурных белка λA, λB и λC кодируются сегментами L гена L1, L2 и L3 соответственно. Сегменты M1 и M2 кодируют два структурных белка (μA и μB) соответственно, в то время как сегмент M3 кодирует неструктурный белок (μNS).Три σ-белка σC, σA и σB кодируются сегментами S1, S2 и S3 соответственно, тогда как сегмент S4 кодирует неструктурный белок σNS. Сегмент S1 кодирует два дополнительных неструктурных белка, а именно p10 и p17 [7, 10].
На основании анализа последовательностей различных генов сообщалось, что реовирусы индейки отличаются от реовирусов кур [20, 44, 45, 52, 88]. Например, сходство нуклеотидной и аминокислотной последовательности сегмента S3, кодирующего сигма-B-белок реовируса индейки (TRV), было обнаружено примерно 61 и 78-80%, соответственно, по сравнению с изолятами куриного реовируса [88].Day et al. [20] сообщили, что белок сигма C гена S1 TRV имеет 57% аминокислотную идентичность с таковой у куриного реовируса (эталонный штамм S1133), тогда как белки p10 и p17 имеют 72 и 61% идентичности, соответственно, с соответствующими белками S1133.
Белок сигма С реовируса млекопитающих и слитый реовирус Нельсона Бэй на 25–28% имеют аминокислотную идентичность с белком сигма С TRV. Ранее мы сообщали, что сходство последовательностей гена S4 на уровне нуклеотидов TRVs было 71.1–74,2% по сравнению с реовирусами куриного происхождения [45]. Эти исследования показывают, что TRV образуют отдельную группу по сравнению с куриными и другими изолятами птичьего реовируса. В результате TRV потенциально можно рассматривать как отдельный вид вируса в подгруппе 2 рода Orthoreovirus . Эта точка зрения дополнительно подтверждается экспериментальными исследованиями, в которых TRV не смогли вызвать признаки заболевания у цыплят, хотя они могут реплицироваться на низких уровнях в кишечном тракте цыплят.Более того, высокопатогенный штамм реовируса куриного происхождения (ARV-1733), вызывающий высокую смертность цыплят, не вызывал клинических признаков у промышленных индеек и имел лишь легкие клинические признаки у индюков, свободных от специфических патогенов [93].
Заболеваемость и распространение
Около 85–90% изолятов реовируса не являются патогенными. Патогенные штаммы реовирусов были связаны с различными заболеваниями кур и индеек. У индеек этот вирус был обнаружен при нескольких различных кишечных заболеваниях, а именно LTS, PEC, PES, миокардите, а недавно и при артрите / теносиновите [6, 28, 39, 44, 47, 68, 69, 90, 91].При исследовании 33 стад индюшат 46% стад оказались положительными на реовирусы [75]. Jindal et al. [44] проверили содержимое кишечника 43 стад, пораженных PES, в 2007–2008 гг. И сообщили о наличии реовируса в 40% стад. Энтерит, вызванный реовирусом, в основном встречается у молодняка индейки в возрасте 1–7 недель, а заболеваемость выше у птиц в возрасте 1–3 недель, которая уменьшается с увеличением возраста.
Реовирусы обнаружены также у практически здоровых кур и индеек [45, 68, 75].При исследовании пяти здоровых родительских стад индюшат 10,4% из 193 протестированных образцов (в возрасте от 1 до 9 недель) были положительными на реовирус [45]. Тестирование образцов кишечника 33 промышленных стад индейки из всех регионов США в течение 2005 и 2006 гг. Выявило присутствие реовируса в 45,5% стад [75].
Передача
Горизонтальная передача фекально-оральным путем является обычным явлением. Из-за стабильности вируса в окружающей среде также может происходить механическая передача.Различные штаммы реовируса различаются по способности распространяться по горизонтали. Подозревается, но не доказано, вертикальная передача кишечной реовирусной инфекции.
Клинические признаки и патология
Пораженные птицы страдают диареей, депрессией, взъерошенными перьями, а также сниженным набором веса [93] и смертностью. Инфекция АРВ может также предрасполагать птиц к вторичным осложнениям или действовать синергетически вместе с другими инфекциями. Сообщалось, что АРВ-инфекция может усиливать кокцидиальную инфекцию у цыплят-бройлеров [85].Также существует возможность синергического действия TRV с другими патогенами у индеек.
В большинстве случаев содержимое кишечника пенистое и водянистое. В некоторых случаях наблюдается атрофия бурсы Фабрициуса [22, 93]. Поражения кишечника состоят из легкой гиперплазии крипт из-за инфильтрации лимфоцитов, гетерофилов и эозинофилов в собственную пластинку и подслизистую оболочку через 11–14 дней после заражения. В сумке Фабрициуса наблюдается умеренное или тяжелое истощение фолликулярных лимфоцитов [22, 93].Помимо атрофии бурсальной сумки, может наблюдаться лимфоидное истощение от легкой до умеренной степени в селезенке и лимфоцитарная инфильтрация в печени, поджелудочной железе, сердце и преджелудках.
Диагноз
Диарея в стаде является признаком кишечной инфекции. Трудно точно определить вирус / возбудитель только на основе диареи, и для подтверждения необходимы лабораторные тесты. Электронная микроскопия может выявить присутствие вирусных частиц размером ~ 70–80 нм в кишечнике инфицированных птиц.Вирус хорошо растет в курином яйце с эмбрионом через желточный мешок или хориоаллантоисную мембрану, а также был выделен в первичных клетках печени куриного эмбриона (CEL), клетках почек куриного эмбриона (CEK) и клетках фибросаркомы японского перепела (QT-35). Из-за фузогенной природы TRV в инокулированных культурах клеток наблюдается характерный цитопатический эффект (ЦПЭ). Для обнаружения антител против реовируса цыплят коммерчески доступны наборы ELISA, но в настоящее время таких специфических наборов для реовируса индейки нет.Недавно мы разработали тест ELISA для обнаружения антител против реовируса индейки. В этом тесте мы клонировали и экспрессировали белок сигма C S1 и использовали его в качестве антигена для обнаружения антител против TRV (неопубликованные данные Mor et al.).
Доступно несколько тестов ОТ-ПЦР, нацеленных на различные генные сегменты реовируса [4, 20, 44, 45, 52, 88]. ОТ-ПЦР в реальном времени — это наиболее чувствительный и специфический тест, с помощью которого можно количественно определить вирусную нагрузку в клинических образцах. Был разработан надежный, сверхчувствительный и точный количественный анализ на АРВ с помощью Light Cycler SYBR Green на основе ОТ-ПЦР в реальном времени, который может обнаруживать 39 копий / мкл геномной РНК АРВ [53].Недавно в нашей лаборатории была разработана высокочувствительная и специфическая ОТ-ПЦР в реальном времени для обнаружения TRV. Мы разработали праймеры и зонды из сегмента генома S4. Тест специфичен для TRV (энтерических вирусов индейки и реовирусов артрита) и может обнаруживать до 10 копий транскрибированной in vitro стандартной вирусной РНК на реакцию (Mor et al. Неопубликованные данные).
Профилактика и контроль
Эффективного лечения реовирусных инфекций не существует. Вирус является повсеместным по своей природе, и из-за его высокой стабильности в окружающей среде трудно избавить птиц и фермы от реовируса.В настоящее время вакцины против реовируса индейки нет, и поэтому биозащита остается единственным способом борьбы с инфекцией.
Ротавирус
Ротавирусы являются важными патогенами, которые вызывают неонатальную диарею у людей, а также у многих животных и птиц. Этот вирус также был обнаружен у коммерческих цыплят-бройлеров, страдающих синдромом короткого роста и задержки роста [72].
Геном
Ротавирусы — это РНК-вирусы, принадлежащие к роду Rotavirus и семейству Reoviridae .Геном ротавируса состоит из 11 сегментов двухцепочечной РНК; РНК окружена тройным слоем икосаэдрического белкового капсида. Каждый сегмент кодирует как минимум один белок. Внешний белковый слой состоит из двух основных нейтрализующих антигенов — вирусного белка 4 (VP4) и вирусного белка 7 (VP7). Вирусный белок 6 (VP6) второго слоя капсида называется групповым антигеном, который используется для обнаружения и классификации ротавирусов на отдельные группы. Кроме того, по крайней мере пять неструктурных белков кодируются геномом ротавируса.
Основываясь на антигенных свойствах VP6, ICTV разделил ротавирусы на пять серологических видов (A – E) и два дополнительных вида (F и G) [81], которые также называются группами ротавирусов. Известно, что группы A, B и C инфицируют людей и животных, а группы D, E, F и G заражают только животных, в основном птиц [50, 63, 102]. Недавно было сообщено о потенциально новом виде RV, ротавирусе H [ 64 ]. Эти авторы утверждают, что значение отсечения аминокислотной последовательности 53% позволяет дифференцировать различные виды ротавирусов.
В отличие от ротавирусов птиц группы A, ротавирусы группы D не могут размножаться в клетках MA104 [ 25 ] и в основном идентифицируются по схеме электрофоретической миграции 5: 2: 2: 2 их сегментов генома, что значительно отличается от паттерна 5: 1: 3: 2 ротавирусов птичьей группы А. Однако классификация штамма ротавируса только на основе его профиля миграции РНК является проблематичной из-за возможного возникновения перестройки генома. В отличие от ротавирусов группы А, которые преимущественно инфицируют клетки двенадцатиперстной кишки, вирусы группы D имеют склонность к тощей и подвздошной кишке.
Ротавирус индейского происхождения отличается от ротавируса куриного происхождения [58]. Например, сообщалось, что гомология аминокислотной последовательности гена VP7 штамма Ty-1 (индейка) и гена Ch-2 VP7 птиц составляет только 70% [58]. Вариабельные эпитопные области A, B и C Ty-1 были уникальными по сравнению с таковыми у Ch-2 и других штаммов, представляющих серотипы 14G. Низкая гомология (53%) областей A и C Ty-1 и Ch-2 предполагает, что штамм Ty-1 может иметь другой серотип, чем эталонный штамм G7 Ch-2.Сравнение последовательностей различных генов (VP4, VP6, VP7, NSP5) ротавирусов индейки и кур показало, что среди ротавирусов птичьей группы А может происходить межвидовая передача и реассортация [87].
Заболеваемость и распространение
Ротавирусы распространены по всему миру и были зарегистрированы как у здоровых, так и у пораженных энтеритом индеек [44, 45, 69, 71, 75, 77, 99]. Периодический мониторинг товарных стад индейки выявил циркуляцию четырех групп (на основе анализа последовательности гена NSP4) ротавирусов.Среди четырех групп ротавирусов было 96,1–97,5 и 97,5–99,55% сходства на уровне нуклеотидов и аминокислот соответственно [77]. Ротавирусы присутствовали до размещения, а также после размещения до 12-недельного возраста [77]. В другом исследовании, в котором участвовали 33 стада индейки и 43 стада кур, ротавирусы были обнаружены в 46% стадах цыплят и 70% стадах индейки [75]. Мы также сообщали о присутствии ротавирусов в практически здоровых стадах производителей индейки с максимальной встречаемостью до 5-недельного возраста [45].В другом исследовании 93% стад, пораженных PES, были положительными на ротавирусы [44]. В контрольном исследовании Mor et al. [68] сообщили, что шесть (7,5%) пулов образцов фекалий были положительными на ротавирус в стадах, пораженных LTS, в то время как 13 (33%) пулов образцов были положительными из стада без LTS.
Выявление ротавирусов выше у молодых цыплят, и уровень обнаружения снижается с возрастом. При наблюдении PES и LTS максимальное количество цыплят было положительным на ротавирус в возрасте 2–5 недель с последующим уменьшением количества положительных образцов в возрасте 8–9 недель [44, 45, 68].Было показано, что праймеры на основе высококонсервативной области генома ротавируса (в частности, неструктурных белков NSP3 или NSP4) подходят для идентификации ротавируса в различных клинических образцах, образцах животных или окружающей среды с помощью обычных или в реальном времени RT- ПЦР [51, 84]. В исследовании, проведенном в Европе, ротавирусы кур и индеек были идентифицированы с использованием двух разных методов: электрофореза в полиакриламидном геле с ОТ-ПЦР и анализов ОТ-ПЦР в реальном времени, специфичных для ротавирусов групп A и D [71].Эти результаты показывают, что необходимы дальнейшие исследования для точного определения типов ротавирусов, циркулирующих в стадах индейки в различных географических точках.
Передача
Фекальный оральный путь является основным путем горизонтальной передачи. Вирус может выжить в окружающей среде в течение длительного времени, и, следовательно, механическая передача может играть роль в его передаче. Есть подозрение на вертикальную передачу ротавируса, но это пока не подтверждено. Например, Pantin-Jackwood et al.[77] обнаружили ротавирусы в 50% проб, исследованных до помещения птицы. Основываясь на этой информации, авторы предположили, что ротавирус может передаваться вертикально. Мы также обнаружили ротавирусы у 2-дневных коммерческих и родительских домашних птиц [45, 47], что указывает на возможность вертикальной передачи ротавируса.
Также описана межвидовая передача ротавируса [87, 103]. В одном исследовании ротавирусы крупного рогатого скота группы А (n = 6) были обнаружены в кишечном содержимом индюков [2].Электроферотипы показали картину миграции, идентичную вирусу диареи телят в Небраске, а полная филогения гена NSP4 показала, что все шесть штаммов сегрегированы по генотипу E2, что подтверждает их принадлежность к ротавирусу крупного рогатого скота. Ротавирусы птиц не обнаружены у людей, и ротавирусы человека не содержат последовательностей ротавирусов птиц [87], что указывает на низкую вероятность заражения человека ротавирусами птиц.
Клинические признаки и патология
Тупость, депрессия, водянистый помет, потеря аппетита, склеивание вентиляционных отверстий и снижение прибавки в весе — постоянные признаки у пораженных птиц.Смертность может присутствовать или отсутствовать, а заболеваемость может варьироваться. Ротавирусная инфекция кур-несушек может привести к снижению яйценоскости.
В большинстве случаев наблюдается пенистое и водянистое кишечное содержимое. Может наблюдаться бледность кишечного тракта и вздутие слепой кишки с пенистым или непенистым жидким содержимым. Другими крупными патологическими проявлениями являются обезвоживание, задержка роста и пастообразные выделения. Также отмечаются мультифокальные и поверхностные эрозии двенадцатиперстной и тощей кишки. Наиболее частыми микроскопическими изменениями являются гиперцеллюлярность собственной пластинки из-за инфильтрации гетерофилов, эозинофилов и мононуклеарных клеток, а также атрофия ворсинок.Сканирующая электронная микроскопия показывает шероховатость поверхности ворсинок, нарушение нормальной морфологии ворсинок и потерю микроворсинок в клетках, расположенных на кончиках ворсинок. У цыплят разные штаммы ротавирусов могут иметь разные предпочтительные сайты репликации. Например, McNulty et al. [66] сообщили, что у экспериментально инфицированных кур ротавирус группы A имел сродство к двенадцатиперстной кишке, тогда как ротавирус группы D предпочитал тощую кишку и подвздошную кишку.
Диагноз
Диарея, обезвоживание и снижение веса стада свидетельствуют о кишечной инфекции.ЭМ кишечного содержимого и фекалий выявляет присутствие вирусных частиц размером ~ 70 нм (приблизительно 50–70 нм) с характерным «колесом» внешнего вида. Несмотря на то, что вирус трудно изолировать, он все же растет в клетках MA104 при обработке трипсином. Тест коагглютинации использовался для обнаружения ротавирусов у индеек и оказался простым и быстрым скрининговым тестом. Электрофорез в полиакриламидном геле обычно используется для определения характерной картины миграции 11 сегментов генома ротавируса и почти так же чувствителен, как ЕМ.
Для обнаружения вируса доступно несколько ОТ-ПЦР и ОТ-ПЦР в реальном времени, нацеленных на различные сегменты генома ротавируса [8, 21, 77]. Недавно было сообщено о высокочувствительной и специфической одностадийной ОТ-ПЦР в реальном времени, нацеленной на неструктурный белок 4 (NSP4), с внутренней системой контроля для обнаружения ротавирусов индейки [1].
Профилактика и борьба
Специальных мер борьбы с ротавирусными инфекциями не существует. Получение цыплят из источников, свободных от ротавирусов, и принятие надлежащих мер биобезопасности — это способы минимизировать заражение ротавирусом в стаде.
Астровирус
Астровирус индейки (TAstV) был впервые обнаружен у индюшат в возрасте 6–11 дней с диареей и повышенной смертностью в Великобритании. В США TAstV был обнаружен у индюшат и позже был назван TAstV-1. Впоследствии TAstV, связанный с PEMS, был выделен и охарактеризован. Этот вирус генетически и иммунологически отличался от ранее описанного изолята США, TAstV-1, и был назван TAstV-2 [56, 86]. Помимо TAstV-1 и TAstV-2, астровирус куриного происхождения i.е. вирус птичьего нефрита (ВНВ) обнаружен также у индеек [26, 75]. ANV сначала был описан как небольшой круглый вирус, затем как пикорнавирус, а позже был определен как астровирус [40]. Таким образом, у индеек обнаружено три типа астровирусов (TAstV-1, TAstV-2 и ANV). Однако данные о последовательностях предполагают, что в природе может присутствовать гораздо больше типов [94]. В 1980-х годах все обнаруженные астровирусы были строго ограничены желудочно-кишечным трактом домашних птиц, и не было никаких признаков системного заболевания.Однако TAstV также был обнаружен в тимусе и сумке, что указывает на возможное влияние на иммунную систему [86]. Сообщалось также об обнаружении астровирусов у кур и индюшат, страдающих кишечными и / или опорно-двигательными расстройствами (теносиновит) [26, 82].
Геном
Астровирусы — это РНК-вирусы без оболочки диаметром 25–35 нм, принадлежащие к роду Avastrovirus семейства Astroviridae . Геном астровируса состоит из 6,8–7 РНК.9 kb и имеет 5′-UTR, за которым следуют три ORF, 3′-UTR и поли-A-хвост. ORF-1a кодирует сериновую протеазу и другие неструктурные белки, ORF-1b для РНК-зависимой РНК-полимеразы и ORF-2 для капсидного белка. Общая длина геномов трех астровирусов, а именно. TAstV-1 (7003 нт), TAstV-2 (7325 нт) и ANV (6927 нт) варьируются; эта длина не включает хвост поли-А [55].
Большинство исследований по обнаружению и характеристике TAstV было проведено молекулярными методами с использованием праймеров, специфичных для гена капсида или полимеразы.Ряд исследований показал вариации на уровне нуклеотидов и аминокислот в гене капсида и / или гене полимеразы TAstVs [13, 44–46, 78, 79, 97]. Сообщается о высоком уровне генетической изменчивости среди астровирусов индейки из стад индейки в США [78]. Идентичность нуклеотидной последовательности в гене капсида, связанная с серотипом и вирусным патогенезом, была всего лишь 69%, но ген полимеразы был более консервативным с 86–99% идентичностью нуклеотидов. Недавно филогенетический анализ белка-предшественника капсида выявил обширные вариации в аминокислотных последовательностях TAstV-2 (81.5–100%), но не в TAstV-1 (96,2–100%) [79]. Мы также обнаружили вариации на уровне аминокислот в гене капсида TAstV-2 [44, 46]. Эти изменения предполагают, что в природе могут существовать различные серотипы астровируса индейки. Для лучшего понимания серотипов TAstV, циркулирующих у индеек, необходимы исследования полных последовательностей генов капсида из явно здоровых и пораженных PEC стад из разных географических регионов. Изменения гена капсида из-за мутаций или рекомбинации могут влиять на антигенность и патогенность вирусов и, следовательно, иметь практическое значение для методов обнаружения вирусов, эпидемиологических исследований и разработки потенциальных вакцин против астровирусных инфекций.
Заболеваемость и распространение
Был проведен ряд исследований для выявления присутствия TAstV в стадах индейки с использованием различных методов. Ранее вирусы в образцах кишечника выявлялись с помощью прямой или иммунной ЭМ. Однако в большинстве недавних исследований используются молекулярные инструменты, такие как ОТ-ПЦР и секвенирование, для обнаружения различных астровирусов [36, 56, 57, 108]. В ходе исследования восьми промышленных предприятий по выращиванию индейки и одного исследовательского подразделения в США, 89,5% из 96 образцов содержимого кишечника были положительными на астровирус с помощью ОТ-ПЦР [77].В исследовании, проведенном в Миннесоте, 84% из 43 стай индеек, пораженных PES, оказались положительными на TAstV-2 с помощью ОТ-ПЦР [44]. Эти образцы не тестировались на наличие TAstV-1 и ANV, поэтому нельзя исключать присутствие TAstV-1 и ANV в этих образцах. Ряд других исследований также выявил высокую распространенность TAstV в стадах индейки в США [75, 77, 78].
Исследования распространенности TAstV также проводились в других странах, где выращивают индюшат [17, 27, 62, 70, 74]. Да Силва и др.[17] сообщили о TAsV у молодых цыплят, пораженных PEC в Бразилии. В другом исследовании, проведенном в Бразилии, TAsV-1, TAstV-2 и ANV были идентифицированы в 64,5, 44,7 и 35,5% образцов кишечника соответственно [70]. Из них TAstV-1 и TAstV-2 были более распространены в стадах индеек, пораженных энтеритом. Domanska-Blicharz et al. [27] сообщили, что TAstV-2 является наиболее распространенным вирусом в Польше; 30 из 77 стад индейки (38,9%) были положительными. TAstV-1 был обнаружен только в девяти стадах, тогда как ANV — в одном стаде. Эти три вируса были обнаружены либо как одиночные, либо как смешанные инфекции.Тестирование 23 образцов кишечника от откормленных индюков с клиническими признаками энтерита выявило присутствие TAstV-2 в 17 образцах в Хорватии [62].
TAstV были обнаружены не только в стадах индюшат, пораженных энтеритом, но также и в практически здоровых стадах индюшат [45, 68, 78]. Jindal et al. [45] протестировали 193 объединенных образца фекалий практически здоровых индюшат-производителей с помощью ОТ-ПЦР и обнаружили, что 47,2% оказались положительными на TAstV-2.
Передача
Передача астровирусов фекально-оральным путем.Здоровые птицы могут подвергаться воздействию астровируса либо при прямом контакте с инфицированными фекалиями, либо с материалами, загрязненными фекалиями. Считается, что астровирусные инфекции видоспецифичны, но недавние сообщения указывают на межвидовую передачу [23, 101]. Так, TAstV-2 и ANV были обнаружены у уток ( Anas platyrhynchos domesticus ) [9]. Межвидовая передача и адаптация вируса к новым хозяевам или совместное инфицирование одного и того же хозяина разными астровирусами может привести к появлению новых астровирусов, которые инфицируют животных или обладают зоонозным потенциалом [24].
Клинические данные и патология
Как и при рота- и реовирусных инфекциях, астровирусы также вызывают диарею, обезвоживание и задержку роста. Сообщалось о подобных наблюдениях диареи и задержки роста у экспериментальных цыплят, инокулированных материалом, содержащим TAstV [48, 49, 76]. Были предприняты экспериментальные исследования, чтобы установить тип клинического заболевания, вызываемого TAstV, имеющими вариантные капсиды, или те, которые различались по источнику происхождения. Например, Pantin-Jackwood et al.[76] изучали патогенность трех различных вирусов TAstV-2 у индюков, свободных от специфических патогенов, и обнаружили, что вирусы TAstV-2 с вариантными капсидами вызывают подобное кишечное заболевание у молодых индеек. Mor et al. [67] обнаружили, что TAstV-2 от птиц PES был более патогенным, чем TAstV-2 от практически здоровых домашних птиц. Астровирус может потенциально нарушать иммунный ответ, тем самым повышая восприимчивость индеек к вторичным бактериальным инфекциям [80].
Обезвоживание, вздутие кишечника, наполненное водянистым содержимым и непереваренной пищей, и расширенная слепая кишка с пенистым содержимым — это результаты вскрытия.Может возникнуть атрофия бурсальной сумки от легкой до умеренной, а также снижение тонуса кишечника. Гистологическими находками являются умеренный некроз эпителия, инфильтраты собственной пластинки и легкая гиперплазия крипт.
Диагноз
Диарея, обезвоживание и снижение веса являются признаками кишечной инфекции. Электронная микроскопия с отрицательным контрастом выявляет вирусные частицы с пятью или шестью выступами на поверхности, напоминающими звезду. Звездообразная морфология не всегда сохраняется или четко различима.По этой причине вирусы AstV от EM можно спутать с пикорнавирусами, пикорнавирусами, энтеровирусами и энтероподобными вирусами. Во-вторых, домашние птицы могут демонстрировать клинические признаки и патологию на более поздней стадии астровирусной инфекции, но в кишечном содержимом может не быть обнаруживаемых астровирусных частиц. В такой ситуации, даже если заболевание возникло, результаты анализов могут быть отрицательными. Молекулярные методы были использованы для обнаружения и дальнейшей характеристики астровирусов. В большинстве анализов на основе ОТ-ПЦР используются праймеры, специфичные для полимеразы или гена капсида TAstV [56, 57, 96].
Смешанные вирусные инфекции
Хотя отдельные вирусы были обнаружены в стадах индейки, пораженных энтеритом, большинство исследований указывают на присутствие двух или более энтеральных вирусов [44, 45, 62, 68, 70, 75, 77, 105, 107 ]. Периодический мониторинг восьми промышленных стад индейки в США выявил постоянное присутствие TAStV и ротавируса [77]. Из 96 протестированных образцов 89,5% были положительными на астровирус и 67,7% на ротавирус. Все стада были отрицательными на TCoV, реовирус и аденовирус группы 1 во всех точках отбора проб.В другом исследовании присутствие астровирусов было зарегистрировано в 100% протестированных стад индейки [75]. TAstV-2 и TAstV-1 были обнаружены в 100 и 15,4% стад индейки соответственно. Кроме того, 12,5% стад индейки были положительными на ANV. Ротавирусы и реовирусы присутствовали в 69,7 и 45,5% стад соответственно, при этом коронавирус и аденовирус не обнаружены.
Кишечное содержимое 43 стад PES в Миннесоте было проверено на наличие энтеральных вирусов методом RT-PCR.Все 43 стада оказались положительными на ротавирус, TAstV-2 или реовирус. Во всех случаях вирусы были обнаружены либо по отдельности, либо в комбинации из 2 или 3. Восемь стай (19%) были положительными на один вирус, а остальные 35 (81%) имели комбинацию вирусов. Пятнадцать стад оказались положительными на все три вируса. Ни одно из протестированных стад не было признано положительным на TCoV [44]. Исследования, проведенные в Бразилии, показали наличие 69,7% множественных вирусов в образцах кишечника индеек. Образцы кишечника чаще всего содержали 3–4 вируса на образец, причем вирусы из семейства астровирусов (TAstV-1, TAstV-2 или ANV, или их комбинация) присутствовали во всех образцах, которые дали положительный результат на вирусы [70 ].
Возможно, что комбинация вирусов может вызвать более серьезные побочные эффекты у домашних птиц, чем отдельные вирусы. В одном исследовании индюшатина, инокулированная комбинацией TAstV-2, реовируса индейки и ротавируса, имела значительно меньшую массу тела, чем контрольная группа [91]. В экспериментальных исследованиях мы также сообщали, что пероральная инокуляция цыплят материалом PES (положительным по ротавирусу, TAstV-2 и Salmonella) приводила к значительной задержке роста на срок до 50 дней после инокуляции, и ни в какой момент после инокуляции не наблюдалось. веса оспариваемых птиц сходятся с весом контрольных.Содержимое кишечника было водянистым и пенистым / пенистым (рис.,) У цыплят, инокулированных материалом PES [49]. Точно так же индюшатины, инокулированные содержимым кишечника птиц, пораженных LTS, имели значительно меньшую массу тела в возрасте 20 недель по сравнению с индюшатинами, инокулированными содержимым кишечника птиц без LTS [68]. Такая степень замедления роста из-за энтерита домашней птицы может привести к значительным экономическим потерям для производителей индейки, поскольку пораженная птица может не достичь ожидаемого веса при продаже.На уровне фермы нельзя исключать роль агентов, отличных от этих четырех распространенных вирусов (TAstV, ротавирус, реовирус и коронавирус), в возникновении и / или повышении серьезности кишечных вирусов. Кроме того, факторы питания и менеджмент также могут играть роль в возникновении энтерита у стада. Такие шаги, как улучшение методов лечения и противомикробная терапия для минимизации потерь, имели ограниченный успех [5]. Вакцины против кишечных вирусов в настоящее время нет.
Водянистое и пенистое содержимое слепой кишки птицы, инокулированной перорально кишечным материалом птиц, пораженных синдромом энтерита домашних птиц
Водянистое и пенистое содержимое слепой кишки птицы, инокулированной перорально кишечным материалом птиц, пораженных синдромом энтерита домашних птиц.Слепая кишка разрезана
Кишечные вирусы обычно обнаруживаются с помощью ЭМ, серологии и ПЦР. Большинство из этих вирусов трудно вырастить в зародышевых яйцах курицы / индейки и клеточных культурах [95]. Таким образом, было разработано несколько анализов одиночной и множественной RT-PCR (mRT-PCR) для обнаружения астровируса индейки, ротавируса индейки, коронавируса индейки и реовируса индейки [21, 43, 56, 77, 89, 92]. Мультиплексные анализы RT-PCR экономят время и деньги, поскольку два или более вируса могут быть обнаружены в одной реакции.
Общие выводы и направления на будущее
Кишечные вирусы (TAstV, ротавирус, реовирус и коронавирус) широко распространены в стадах индейки, но их клиническое значение неясно. Полное влияние этих вирусов на производительность стада требует тщательного изучения. Хотя кишечные заболевания, вызванные вирусами, являются экономическим бременем для фермеров, выращивающих индейки, ни один вирус не появился в качестве вероятного возбудителя, чтобы можно было сформулировать соответствующие меры по профилактике и контролю [18].В нескольких исследованиях одновременно были обнаружены два или более разных вируса, что указывает на то, что несколько вирусов могут участвовать в патогенезе энтерита. Преобладание вирусов одного и того же рода в кишечнике как здоровых, так и плохо функционирующих птиц указывает на существование разных патотипов вируса. Необходимы дальнейшие исследования, чтобы определить обоснованность этих утверждений.
С появлением молекулярных диагностических тестов и секвенирования наше понимание кишечных вирусов расширилось.Также существует вероятность того, что наши текущие методы не могут обнаружить некоторые еще неизвестные вирусы / патогены, которые могут иметь последствия для энтерита индейки. Доступность и использование высокопроизводительных методов пиросеквенирования нуклеиновых кислот нового поколения, таких как секвенирование Illumina, может выявить последовательности агентов, о которых ранее не сообщалось у индеек. Метагеномные исследования РНК-вирусов в кишечнике индеек, страдающих кишечным заболеванием, выявили присутствие последовательностей вирусов дцРНК ( Reoviridae и пикобирнавирусы) и вирусов оцРНК ( Caliciviridae , Leviviridae , Picornaviridae Astroviridae и Picornaviridae ). [18].Обнаружение неизвестных вирусов методом метагеномики проложит путь к разработке методов диагностики этих вирусов. Кроме того, необходимо определить, почему одни и те же типы вируса обнаруживаются в стадах индейки, пораженных энтеритом, а также в явно здоровых стадах. Полное понимание роли кишечных вирусов и других патогенов в кишечных болезнях индеек будет иметь большое значение при разработке профилактических и контрольных мер.
Ссылки
1. Акимкин В., Биндель Ф., Хоферер М., Стинг Р., Полли Б., Ханель А., Хафез Х.М.Одношаговая RT-qPCR с системой внутреннего контроля для обнаружения ротавирусов индейки в образцах фекалий. J Virol Methods. 2011. 177 (1): 112–117. [PubMed] [Google Scholar] 2. Асано К. М., Грегори Ф., Соуза С. П., Ротава Д., Оливейра Р. Н., Вильярреал Л. Я., Рихтценхайн Л. Дж., Брандао ЧП. Ротавирус крупного рогатого скота у индеек с энтеритом. Птичий Дис. 2011; 55 (4): 697–700. [PubMed] [Google Scholar] 3. Трепет О.О., Али А., Элайш М., Ибрагим М., Мурджа М., Пантин-Джеквуд М., Саиф Ю.М., Ли К.В. Влияние коронавирусной инфекции на репродуктивную способность кур индюков.Птичий Дис. 2013; 57: 650–656. [PubMed] [Google Scholar] 4. Баньяи К., Дандар Э., Дорси К.М., Мато Т., Паля В. Геномное созвездие нового штамма ортореовируса птиц, связанного с синдромом низкорослости у бройлеров. Гены вирусов. 2011; 42: 82–89. [PubMed] [Google Scholar]5. Барнс Х. Дж., Гай Дж. С.. Синдром энтерита-смертности птицы. В: Саиф Ю.М., Барнс Х.Дж., Глиссон-младший, Фэдли А.М., Макдугалд Л.Р., Суэйн Д.Е., редакторы. Болезни птицы. 11-е изд. Эймс: Издательство государственного университета Айовы; 2003. С. 1171–1180.
6. Barnes HJ, Guy JS, Vaillancourt JP. Комплекс энтерита птицы. Rev Sci Tech. 2000. 19: 565–588. [PubMed] [Google Scholar] 7. Бенавенте Дж., Мартинес-Костас Дж. Птичий реовирус: структура и биология. Virus Res. 2007. 123: 105–119. [PubMed] [Google Scholar] 8. Безерра Д.А., да Силва Р.Р., Кайано Дж. Х., Сильвестр Р. В., де Соуза Оливейра Д., Линьярес С., Габбай Ю. Б., Маскареньяс Дж. Д.. Обнаружение ротавируса группы D птиц с использованием полимеразной цепной реакции для гена VP6. J Virol Methods. 2012; 185: 189–192. [PubMed] [Google Scholar] 9.Bidin M, Bidin Z, Majnaric D, Tisljar M, Lojkic I. Циркуляция и филогенетическая взаимосвязь астровирусов куриного и индейского происхождения, обнаруженных у домашних уток ( Anas platyrhynchos domesticus ) Avian Pathol. 2012. 41 (6): 555–562. [PubMed] [Google Scholar] 10. Боделон Дж., Лабрада Л., Мартинез-Костас Дж., Бенавенте Дж. Сегмент генома реовируса птиц S1 представляет собой функционально трицистронный ген, который экспрессирует один структурный и два неструктурных белка в инфицированных клетках. Вирусология. 2001; 290: 181–191.[PubMed] [Google Scholar] 11. Бреслин Дж. Дж., Смит Л. Г., Барнс Г. Дж., Гай Дж. С.. Сравнение процедур выделения вируса, иммуногистохимии и полимеразной цепной реакции с обратной транскриптазой для обнаружения коронавируса индейки. Птичий Дис. 2000. 44: 624–631. [PubMed] [Google Scholar] 12. Бангер А. Н., Чакон Дж. Л., Джонс Р. К., Феррейра А. Дж.. Обнаружение и молекулярная характеристика гена 3 и 5 коронавируса индейки у индюшат с тяжелым энтеритом в Бразилии. Птичий Дис. 2009. 53: 356–362. [PubMed] [Google Scholar] 13.Канелли Э., Кордиоли П., Барбьери И., Кателла А., Пеннелли Д., Церути Р., Морено А., Лавацца А. Астровирусы как возбудители энтерита домашней птицы: генетическая характеристика и лонгитюдные исследования в полевых условиях. Птичий Дис. 2012; 56: 173–182. [PubMed] [Google Scholar] 14. Cardoso TC, Castanheira TL, Teixeira MC, Rosa AC, Hirata KY, Astolphi RD, Luvizotto MC. Валидация иммуногистохимического анализа для обнаружения коронавируса индейки: быстрый и простой инструмент для скрининга в условиях ограниченных ресурсов. Poult Sci.2008. 87 (7): 1347–1352. [Бесплатная статья PMC] [PubMed] [Google Scholar] 15. Чен Ю.Н., Ву СС, Брайан Т., Хупер Т., Шредер Д., Лин Т.Л. Специфическая полимеразная цепная реакция обратной транскрипции в режиме реального времени для обнаружения и количественного определения РНК коронавируса индейки в тканях и фекалиях индеек, инфицированных коронавирусом индейки. J Virol Methods. 2010. 163: 452–458. [Бесплатная статья PMC] [PubMed] [Google Scholar] 16. Калвер Ф.А., Бриттон П., Кавана Д. Обнаружение с помощью ОТ-ПЦР птичьих коронавирусов галлиформных птиц (курица, индейка, фазан) и у попугая.Методы Мол биол. 2008; 454: 35–42. [Бесплатная статья PMC] [PubMed] [Google Scholar] 17. Да Силва SEL, Бонетти AM, Petrocelli ATM, Ferrari HF, Luvizotto MCR, Cardoso TC. Обнаружение астровируса индейки у молодняка, пораженного комплексом энтерита домашней птицы, в Бразилии. J Vet Med Sci. 2008. 70 (6): 629–631. [PubMed] [Google Scholar]19. Дэй Дж. М., Гондер Э., Дженнингс С., Ривес Д., Тилли Б., Вуминг Б. Молекулярная характеристика кишечных коронавирусов индейки, циркулирующих в США в 2012 году. Документ, представленный на ежегодном собрании Американской ассоциации патологоанатома в Чикаго, штат Иллинойс.2013.
20. Day JM, Pantin-Jackwood MJ, Spackman E. Последовательный и филогенетический анализ сегмента S1 генома реовирусов индейского происхождения. Гены вирусов. 2007. 35 (2): 235–242. [PubMed] [Google Scholar] 21. Дэй JM, Spackman E, Пантин-Джеквуд MJ. Мультиплексный ОТ-ПЦР-тест для дифференциальной идентификации астровируса индейки типа 1, астровируса индейки типа 2, астровируса цыпленка, вируса птичьего нефрита и птичьего ротавируса. Птичий Дис. 2007. 51: 681–684. [PubMed] [Google Scholar] 22. Дэй JM, Spackman E, Пантин-Джеквуд MJ.Иммунная дисфункция, вызванная реовирусом индейского происхождения, у свободных от конкретных патогенов и промышленных индюшат. Птичий Дис. 2008. 52 (3): 387–391. [PubMed] [Google Scholar] 23. Де Баттисти С., Сальвиато А., Йонассен С.М., Тоффан А., Капуа I, Каттоли Г. Генетическая характеристика астровирусов, обнаруженных у цесарок ( Numida meleagris ), выявляет отдельный генотип и предполагает межвидовую передачу между индейкой и цесаркой. Arch Virol. 2012. 157 (7): 1329–1337. [PubMed] [Google Scholar] 24. Де Бенедиктис П., Шульц-Черри С., Бернем А., Каттоли Г.Астровирусные инфекции у людей и животных — молекулярная биология, генетическое разнообразие и межвидовая передача. Заразить Genet Evol. 2011. 11 (7): 1529–1544. [Бесплатная статья PMC] [PubMed] [Google Scholar] 25. Девитт CM, Рейнольдс DL. Характеристика ротавирусов группы D. Птичий Дис. 1993; 37: 749–755. [PubMed] [Google Scholar] 26. de Wit JJ, ten Dam GB, van de Laar JMAM, Biermann Y, Verstegen I, Edens F, Schrier CC. Обнаружение и характеристика нового астровируса у кур и индеек с кишечными расстройствами и нарушениями опорно-двигательного аппарата.Avian Pathol. 2011; 40: 453–461. [PubMed] [Google Scholar] 27. Доманска-Бличарц К., Серока А., Минта З. Годовое молекулярное исследование астровирусной инфекции у индеек в Польше. Arch Virol. 2011. 156 (6): 1065–1072. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Франка М., Креспо Р., Чин Р., Вулкок П., Шивапрасад Х.Л. Ретроспективное исследование миокардита, ассоциированного с реовирусом, у индеек. Птичий Дис. 2010. 54 (3): 1026–1031. [PubMed] [Google Scholar] 30. Gomaa MH, Yoo D, Ojkic D, Barta JR. Распространенность коронавируса индейки у североамериканских индеек определяется с помощью недавно разработанного иммуноферментного анализа на основе рекомбинантного антигена.Clin Vaccine Immunol. 2008. 15 (12): 1839–1844. [Бесплатная статья PMC] [PubMed] [Google Scholar] 31. Gomaa MH, Yoo D, Ojkic D, Barta JR. Использование рекомбинантного полипептида-шипа S1 для разработки ELISA-анализа антител, специфичных к TCoV. Vet Microbiol. 2009. 138 (3–4): 281–288. [Бесплатная статья PMC] [PubMed] [Google Scholar] 32. Gomaa MH, Yoo D, Ojkic D, Barta JR. Заражение патогенным изолятом коронавируса индейки отрицательно влияет на показатели роста и морфологию кишечника молодняка индейки в Канаде. Avian Pathol.2009. 38: 279–286. [PubMed] [Google Scholar] 33. Гомес Д.Е., Хирата К.Ю., Сахеки К., Роза А.С., Лувизотто М.С., Кардосо ТК. Патология и тканевое распределение коронавируса индейки у экспериментально инфицированных цыплят и индюшат. J Comp Pathol. 2010. 143 (1): 8–13. [Бесплатная статья PMC] [PubMed] [Google Scholar] 34. Гай JS. Коронавирусный энтерит в Турции. В: Саиф Ю.М., Барнс Х.Дж., Глиссон-младший, Фэдли А.М., Макдугалд Л.Р., Суэйн Д.Е., редакторы. Болезнь птицы. 11. Эймс: Издательство государственного университета Айовы; 2003. С. 300–307. [Google Scholar] 35.Гай JS. Коронавирусный энтерит в Турции. В: Саиф YM, Fadly AM, Glisson JR, McDougald LR, Nolan LK, Swayne DE, редакторы. Болезни птицы. 12. Эймс: специалист по издательскому делу Wiley-Blackwell; 2008. С. 330–338. [Google Scholar] 36. Guy JS, Miles AM, Smith L, Fuller FJ, Schultz-Cherry S. Антигенная и геномная характеристика энтеровирусоподобного вируса индейки (Северная Каролина, изолят 1988 г.): идентификация вируса как астровируса индейки 2. Avian Dis. 2004. 48 (1): 206–211. [PubMed] [Google Scholar] 37.Гай Дж. С., Смит Л. Г., Бреслин Дж. Дж., Пакпиньо С. Разработка конкурентного иммуноферментного анализа для обнаружения антител к коронавирусу индейки. Птичий Дис. 2002. 46 (2): 334–341. [PubMed] [Google Scholar] 38. Гай Дж. С., Смит Л. Г., Бреслин Дж. Дж., Вайланкур Дж. П., Барнс Х. Дж. Высокая смертность и подавление роста, экспериментально вызванные у молодых индеек двойным заражением энтеропатогенным вирусом Escherichia coli и коронавирусом индейки. Птичий Дис. 2000. 44: 105–113. [PubMed] [Google Scholar] 39.Heggen-Peay CL, Qureshi MA, Edens FW, Sherry B, Wakenell PS, O’Connell PH, Schat KA. Выделение реовируса от синдрома энтерита и смертности домашних птиц и его патогенность у индюшат. Птичий Дис. 2002. 46 (1): 32–47. [PubMed] [Google Scholar] 40. Имада Т., Ямагути С., Масе М, Цукамото К., Кубо М., Морока А. Вирус птичьего нефрита (ANV) как новый член семейства Astroviridae и конструирование инфекционной кДНК ANV. J Virol. 2000; 74: 8487–8493. [Бесплатная статья PMC] [PubMed] [Google Scholar] 41.Исмаил М.М., Тан А.Ю., Саиф Ю.М. Патогенность коронавируса индейки у индеек и кур. Птичий Дис. 2003. 47: 515–522. [PubMed] [Google Scholar] 42. Джеквуд М.У., Бойнтон Т.О., Хилт Д.А., Маккинли Е.Т., Киссинджер Дж.С., Патерсон А.Х., Робертсон Дж., Лемке С., МакКолл А.В., Уильямс С.М., Джеквуд Дж. В., Берд Л.А. Возникновение коронавируса группы 3 в результате рекомбинации. Вирусология. 2010; 398: 98–108. [Бесплатная статья PMC] [PubMed] [Google Scholar] 43. Джиндал Н., Чандер И., Патнаяк Д.П., Мор СК, Зиглер А.Ф., Гоял С.М. Мультиплексная RT-PCR для обнаружения астровирусов, ротавирусов и реовирусов у индеек.Птичий Дис. 2012; 56: 592–596. [PubMed] [Google Scholar] 44. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М. Выявление и молекулярная характеристика кишечных вирусов синдрома энтерита домашней птицы у индеек. Poult Sci. 2010. 89: 217–226. [Бесплатная статья PMC] [PubMed] [Google Scholar] 45. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М. Обнаружение и молекулярная характеристика кишечных вирусов у племенных индюков. Avian Pathol. 2010; 39: 53–61. [PubMed] [Google Scholar] 46. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М.Сравнение последовательностей капсидных генов астровируса-2 индейки у индюшат, пораженных синдромом энтерита, и у практически здоровых индюшат. Arch Virol. 2011; 156: 969–977. [PubMed] [Google Scholar] 47. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Ретроспективное исследование синдрома энтерита домашней птицы в Миннесоте. Птичий Дис. 2009; 53: 268–275. [PubMed] [Google Scholar] 48. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Экспериментальное воспроизведение синдрома энтерита домашней птицы: клинические данные, реакция роста и микробиология.Poult Sci. 2009; 88: 949–958. [Бесплатная статья PMC] [PubMed] [Google Scholar] 49. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Продолжительность угнетения роста и выделения патогенов при синдроме энтерита экспериментально воспроизведенных индюшат. Птичий Дис. 2009; 53: 517–522. [PubMed] [Google Scholar] 50. Johne R, Otto P, Roth B, Löhren U, Belnap D, Reetz J, Trojnar E. Анализ последовательности сегмента генома, кодирующего VP6, ротавирусов птичьей группы F и G. Вирусология. 2011; 412: 384–391. [PubMed] [Google Scholar] 51.Джотикумар Н, Канг Г., Хилл ВР. Широкореактивный анализ TaqMan для обнаружения ротавируса в режиме реального времени с помощью ОТ-ПЦР в клинических образцах и образцах окружающей среды. J Virol Methods. 2009. 155: 126–131. [PubMed] [Google Scholar] 52. Капчинский Д.Р., Селлерс Х.С., Симмонс В., Шульц-Черри С. Анализ последовательности гена S3 из реовируса индейки. Гены вирусов. 2002. 25 (1): 95–100. [Бесплатная статья PMC] [PubMed] [Google Scholar] 53. Кэ GM, Cheng HL, Ke LY, Ji WT, Chulu JL, Liao MH, Chang TJ, Liu HJ. Разработка количественного светового циклера RT-PCR в реальном времени для обнаружения птичьего реовируса.J Virol Methods. 2006; 133 (1): 6–13. [PubMed] [Google Scholar] 54. King AMQ, Lefkowiz E, Adams M, Carstens E. Таксономия вирусов: девятый доклад международного комитета по таксономии вирусов. Уолтем: Academic Press; 2012. [Google Scholar] 55. Koci MD, Schultz-Cherry S. Птичьи астровирусы. Avian Pathol. 2002; 31: 213–227. [PubMed] [Google Scholar] 56. Koci MD, Seal BS, Schultz-Cherry S. Разработка диагностического теста RT-PCR для птичьего астровируса. J Virol Methods. 2000. 90: 79–83. [Бесплатная статья PMC] [PubMed] [Google Scholar] 58.Кул Д.А., Холмс И.Х. Нуклеотидная и расчетная аминокислотные последовательности Ty-1 Vp7 ротавируса птиц значительно отличаются от таковых ротавируса Ch-2. Arch Virol. 1993. 129 (1–4): 227–234. [PubMed] [Google Scholar] 59. Lin TL, Loa CC, Tsai SC, Wu CC, Bryan TA, Thacker HL, Hooper T., Schrader D. Характеристика коронавируса индейки у индюшат с острым энтеритом. Vet Microbiol. 2002. 84 (1–2): 179–186. [Бесплатная статья PMC] [PubMed] [Google Scholar] 60. Линь Т.Л., Лоа СС, Ву СС. Полные последовательности 3′-концевой кодирующей области для генов структурных белков коронавируса индейки.Virus Res. 2004. 106 (1): 61–70. [Бесплатная статья PMC] [PubMed] [Google Scholar] 61. Loa CC, Wu CC, Lin TL. Сравнение 3′-концевых кодирующих областей изолятов коронавируса индейки из Индианы, Северной Каролины и Миннесоты со штаммами коронавируса инфекционного бронхита кур. Интервирология. 2006. 49 (4): 230–238. [Бесплатная статья PMC] [PubMed] [Google Scholar] 62. Lojkic I, Bidin M, Bidin Z, Mikec M. Вирусные агенты, связанные с энтеритом домашней птицы в хорватских промышленных стадах индейки. Acta Vet BRNO. 2010; 79: 91–98.[Google Scholar] 63. Маттейнссенс Дж., Мартелла В., Ван Ранст М. Оценка приоритетной статьи: геномная эволюция, межвидовой барьер хозяина, реассортировка и классификация ротавирусов. Будущий Virol. 2010. 5: 385–390. [Google Scholar] 64. Matthijnssens J, Otto PH, Ciarlet M, Desselberger U, Van Ranst M, Johne R. Значения отсечения на основе последовательности VP6 в качестве критерия для демаркации видов ротавирусов. Arch Virol. 2012. 157 (6): 1177–1182. [PubMed] [Google Scholar] 65. Maurel S, Toquin D, Briand FX, Queguiner M, Allee C, Bertin J, Ravillion L, Retaux C, Turblin V, Morvan H, Eterradossi N.Первые полноразмерные последовательности гена S европейских изолятов выявили дальнейшее разнообразие коронавирусов индейки. Avian Pathol. 2011; 40: 179–189. [PubMed] [Google Scholar] 66. Макналти М.С., Аллан Г.М., Маккракен Р.М. Экспериментальное заражение цыплят ротавирусами: клинические и вирусологические данные. Avian Pathol. 1983; 12: 45–54. [PubMed] [Google Scholar] 67. Мор С.К., Абин М., Коста Дж., Дуррани А., Джиндал Н., Гоял С.М., Патнаяк Д.П. Роль астровируса индейки 2 типа в синдроме энтерита домашних птиц. Poult Sci.2011; 90: 2747–2752. [Бесплатная статья PMC] [PubMed] [Google Scholar] 68. Мор С.К., Шарафельдин Т.А., Абин М., Кромм М., Портер Р.Э., Гоял С.М., Патнаяк Д.П. Возникновение кишечных вирусов при синдроме легкой индейки. Avian Pathol. 2013. 42 (5): 497–501. [PubMed] [Google Scholar] 69. Мор С.К., Шарафельдин Т.А., Портер Р.Э., Зиглер А., Патнаяк Д.П., Гоял С.М. Выделение и характеристика реовируса артрита индейки. Птичий Дис. 2013. 57 (1): 97–103. [PubMed] [Google Scholar] 70. Моура-Альварес Дж., Чакон СП, Сканавини Л.С., Нуньес ЛФН, Астольфи-Феррейра К.С., Джонс Р.К., Пиантино Феррейра А.Дж.Кишечные вирусы в стадах бразильских индейок: частота единичных и множественных вирусных инфекций в зависимости от возраста и клинических признаков кишечного заболевания. Poult Sci. 2013; 92: 945–955. [Бесплатная статья PMC] [PubMed] [Google Scholar] 71. Otto PH, Ahmed MU, Hotzel H, Machnowska P, Reetz J, Roth B, Trojnar E, Johne R. Обнаружение птичьих ротавирусов групп A, D, F и G у больных кур и индеек из Европы и Бангладеш. Vet Microbiol. 2012; 156 (1-2): 8-15. [Бесплатная статья PMC] [PubMed] [Google Scholar] 72.Отто П., Либлер-Тенорио Е.М., Эльшнер М., Ритц Дж., Лорен У., Диллер Р. Обнаружение ротавирусов и поражений кишечника у цыплят-бройлеров в стадах с синдромом короткого роста и задержки роста (RSS) Avian Dis. 2006; 50: 411–418. [PubMed] [Google Scholar] 73. Пакпиньо С., Лей Д.Х., Барнс Х.Дж., Вайланкур Дж. П., Гай Дж. С.. Распространенность энтеропатогенной Escherichia coli в естественных случаях синдрома энтерита-смертности домашних птиц. Птичий Дис. 2002. 46 (2): 360–369. [PubMed] [Google Scholar] 74. Palade EA, Demeter Z, Hornyak A, Nemes C, Kisary J, Rusvai M.Высокая распространенность парвовируса индейки в стадах индейки из Венгрии, страдающих синдромами кишечных заболеваний. Птичий Дис. 2011; 55 (3): 468–475. [PubMed] [Google Scholar] 75. Пантин-Джеквуд MJ, Day JM, Jackwood MW, Spackman E. Кишечные вирусы, обнаруженные молекулярными методами в коммерческих стадах кур и индейок в Соединенных Штатах в период с 2005 по 2006 гг. Птичий Дис. 2008. 52: 235–244. [PubMed] [Google Scholar] 76. Пантин-Джеквуд MJ, Spackman E, Day JM. Патогенез астровирусов индейки 2 типа с вариантными генами капсида у 2-дневных цыплят, свободных от специфических патогенов.Avian Pathol. 2008; 37: 193–201. [PubMed] [Google Scholar] 77. Пантин-Джеквуд MJ, Spackman E, Day JM, Rives D. Периодический мониторинг промышленных индеек на предмет энтеросолюбии указывает на постоянное присутствие астровируса и ротавируса на фермах. Птичий Дис. 2007. 51: 674–680. [PubMed] [Google Scholar] 78. Пантин-Джеквуд MJ, Spackman E, Woolcock PR. Молекулярная характеристика и типирование астровирусов кур и индейки, циркулирующих в США: значение для диагностики. Птичий Дис. 2006; 50: 397–404.[PubMed] [Google Scholar] 79. Пантин-Джеквуд MJ, Strother KO, Mundt E, Zsak L, Day JM, Spackman E. Молекулярная характеристика птичьих астровирусов. Arch Virol. 2011; 156: 235–244. [PubMed] [Google Scholar] 80. Куреши М.А., Саиф Ю.М., Хегген-Пи К.Л., Эденс Ф.В., Хавенштейн ГБ. Индукция функциональных дефектов в макрофагах астровирусом индейки, ассоциированным с энтеритом и синдромом смертности домашних птиц. Птичий Дис. 2001. 45 (4): 853–861. [PubMed] [Google Scholar] 81. Рамиг РФ, Ciarlet M, Mertens PPC, Dermody TS. Ротавирус.В: Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA, редакторы. Таксономия вирусов: восьмой отчет международного комитета по таксономии вирусов. Амстердам: Academic Press; 2005. С. 484–496. [Google Scholar] 82. Рейнольдс Д.Л., Саиф Ю.М. Астровирус: причина кишечного заболевания у индюшат. Птичий Дис. 1986; 30: 728–735. [PubMed] [Google Scholar]83. Роббинс К.М. Эпидемиология и диагностика коронавируса в Турции. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс.2013.
84. Родригес-Диас Дж., Рубилар-Абреу Э., Шпицнер М., Хедлунд К.О., Липранди Ф., Свенссон Л. Дизайн мультиплексной вложенной ПЦР для генотипирования NSP4 ротавируса группы А. J Virol Methods. 2008. 149: 240–245. [PubMed] [Google Scholar] 85. Ruff MD, Rosenberger JK. Сопутствующие инфекции реовирусами и кокцидиями у бройлеров. Птичий Дис. 1985. 29 (2): 465–478. [PubMed] [Google Scholar] 86. Шульц-Черри С., Капчинский Д.Р., Симмонс В.М., Кочи М.Д., Браун С., Барнс Х.Дж. Идентификация агента (ов), связанного с синдромом смертности от энтерита домашней птицы: важность тимуса.Птичий Дис. 2000. 44: 256–265. [PubMed] [Google Scholar] 87. Шуман Т., Хотцель Х., Отто П., Джон Р. Доказательства межвидовой передачи и реассортации среди ротавирусов птичьей группы А. Вирусология. 2009. 386 (2): 334–343. [PubMed] [Google Scholar] 88. Продавцы HS, Linnemann EG, Pereira L, Kapczynski DR. Филогенетический анализ гена белка сигма 2 реовирусов индейки. Птичий Дис. 2004. 48 (3): 651–657. [PubMed] [Google Scholar] 89. Продавцы HS, Koci MD, Linnemann E, Kelley LA, Schultz-Cherry S. Разработка диагностического теста мультиплексной полимеразной цепной реакции с обратной транскрипцией, специфичного для астровируса и коронавируса индейки.Птичий Дис. 2004. 48: 531–539. [PubMed] [Google Scholar] 90. Shivaprasad HL, Franca M, Woolcock PR, Nordhausen R, Day JM, Pantin-Jackwood M. Миокардит, связанный с реовирусом у индюшат. Птичий Дис. 2009. 53: 523–532. [PubMed] [Google Scholar] 91. Spackman E, Day JM, Pantin-Jackwood MJ. Сопутствующие инфекции астровируса, реовируса и ротавируса вызывают снижение прибавки в весе у широкогрудых белых цыплят. Птичий Дис. 2010; 54: 16–21. [PubMed] [Google Scholar] 92. Спакман Э., Капчинский Д., Селлерс Х. Мультиплексная полимеразная цепная реакция обратной транскрипции в реальном времени для обнаружения трех вирусов, связанных с комплексом энтерита домашней птицы: астровируса индейки, коронавируса индейки и реовируса индейки.Птичий Дис. 2005; 49: 86–91. [PubMed] [Google Scholar] 93. Спакман Э., Пантин-Джеквуд М., Дэй Дж. М., Селлерс Х. Патогенез реовирусов индейского происхождения у индеек и кур. Avian Pathol. 2005; 34: 291–296. [PubMed] [Google Scholar] 94. Штамм E, Kelley LA, Schultz-Cherry S, Muse SV, Koci MD. Геномный анализ близкородственных астровирусов. J Virol. 2008. 82: 5099–5103. [Бесплатная статья PMC] [PubMed] [Google Scholar] 95. Сугияма М.К., Гото К., Уэмукай Х., Мори Й., Ито Н., Минамото Н. Присоединение и инфицирование к клеткам MA104 ротавирусов птиц требует присутствия сиаловой кислоты на поверхности клетки.J Vet Med Sci. 2004. 66: 461–463. [PubMed] [Google Scholar] 96. Тан Й., Исмаил М.М., Саиф Ю.М. Разработка иммуноферментного анализа с захватом антигена и ОТ-ПЦР для обнаружения астровирусов индейки. Птичий Дис. 2005. 49 (2): 182–188. [PubMed] [Google Scholar] 97. Тан Й., Мурджа А.М., Саиф Ю.М. Молекулярная характеристика гена капсида двух серотипов астровирусов индейки. Птичий Дис. 2005. 49 (4): 514–519. [PubMed] [Google Scholar] 98. MC Teixeira, MC Luvizotto, Ferrari HF, Mendes AR, da Silva SE, Cardoso TC.Обнаружение коронавируса индейки у коммерческих индюшат в Бразилии. Avian Pathol. 2007. 36 (1): 29–33. [PubMed] [Google Scholar] 99. Тейл К.В., Саиф Ю.М. Возрастные инфекции ротавируса, ротавирусоподобного вируса и атипичного ротавируса в стадах индейки. J Clin Microbiol. 1987. 25 (2): 333–337. [Бесплатная статья PMC] [PubMed] [Google Scholar]100. Тилли Б.Дж., Гондер Э., Мейсон С., Уикс Дж. Вспышка коронавируса в Турции в Северной Каролине. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс.2013.
101. Тоффан А., Катания С., Сальвиато А., Де Баттисти С., Васкеллари М., Тосон М., Капуа I, Каттоли Г. Экспериментальное заражение домашних птиц и цесарок генетически отличными птичьими астровирусами. Avian Pathol. 2012; 41: 429–435. [PubMed] [Google Scholar] 102. Trojnar E, Otto P, Roth B, Reetz J, Johne R. Сегменты генома ротавируса группы D имеют консервативные концы группы A, но кодируют специфичные для группы белки. J Virol. 2010; 84: 10254–10265. [Бесплатная статья PMC] [PubMed] [Google Scholar] 103.Trojnar E, Sachsenroder J, Twardziok S, Reetz J, Otto PH, Johne R. Идентификация ротавируса птичьей группы А, содержащего новый ген VP4, близкий родству к генам ротавирусов млекопитающих. J Gen Virol. 2013; 94: 136–142. [PubMed] [Google Scholar] 104. Велаюдхан Б.Т., Шин Х.Д., Лопес В.К., Хупер Т., Халворсон Д.А., Нагараджа К.В. Анализ обратной транскриптазы-полимеразной цепной реакции для диагностики коронавирусной инфекции индейки. J Vet Diagn Invest. 2003. 15 (6): 592–596. [PubMed] [Google Scholar] 105.Woolcock PR, Shivaprasad HL. Электронно-микроскопическая идентификация вирусов, связанных с энтеритом домашней птицы, у индеек, выращенных в Калифорнии в 1993–2003 гг. Птичий Дис. 2008. 52: 209–213. [PubMed] [Google Scholar]106. Вуминг Б. Вспышка коронавируса на северо-западе Арканзаса. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс. 2013.
107. Ю М., Исмаил М. М., Куреши М. А., Дерт Р. Н., Барнс Х. Дж., Саиф Ю. М.. Вирусные агенты, связанные с энтеритом и синдромом смертности домашних птиц: роль небольшого круглого вируса и коронавируса индейки.Птичий Дис. 2000. 44 (2): 297–304. [PubMed] [Google Scholar] 108. Ю М, Тан Й, Го М, Чжан Кью, Саиф Ю. Характеристика небольшого круглого вируса, связанного с синдромом энтерита и смертности цыплят. Птичий Дис. 2000. 44 (3): 600–610. [PubMed] [Google Scholar] 109. Zsak L, Strother KO, Kisary J. Анализ частичной последовательности генома парвовирусов, связанных с кишечным заболеванием у домашней птицы. Avian Pathol. 2008. 37 (4): 435–441. [PubMed] [Google Scholar]Кишечных вирусов энтерита индейки
Вирусная болезнь.2014 июн; 25 (2): 173–185.
, , иНареш Джиндал
Департамент ветеринарного здравоохранения и эпидемиологии, Колледж ветеринарных наук, Университет ветеринарии и зоотехники Лала Ладжпат Рай, Хисар, 125 004 Индия
9004, Индия. К. МорКафедра ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Сент-Пол, Миннесота 55108 США
Сагар М.Гоял
Департамент ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Сент-Пол, Миннесота 55108 США
Департамент ветеринарного здравоохранения и эпидемиологии, Колледж ветеринарных наук, Ветеринарный университет Лала Ладжпат Рай. Науки о животных, Хисар, 125 004 Индия
Департамент ветеринарной народной медицины Колледж ветеринарной медицины, Университет Миннесоты, Сент-Пол, Миннесота 55108 США
Автор, отвечающий за переписку.Поступило 2 октября 2013 г .; Принято 27 января 2014 г.
Эта статья цитируется в других статьях в PMC.Abstract
Здоровье кишечника очень важно для получения максимальной отдачи с точки зрения увеличения веса и яйценоскости. Кишечные заболевания, такие как комплекс энтерита домашней птицы (PEC) у индеек, не позволяют максимально реализовать их производственный потенциал. Был вовлечен ряд вирусов, бактерий и простейших, но первичная этиология окончательно не установлена. Раньше электронная микроскопия использовалась для обнаружения кишечных вирусов, которые определялись исключительно на основе их морфологии.С появлением методов быстрой молекулярной диагностики и секвенирования нуклеиновых кислот нового поколения исследователи добились больших успехов в идентификации и характеристике вирусов, связанных с PEC. Молекулярные методы также помогли нам идентифицировать патогены, которые ранее не были известны. Региональные и национальные исследования выявили наличие нескольких различных кишечных вирусов в PEC, включая ротавирус, астровирус, реовирус и коронавирус, по отдельности или в комбинации.По-прежнему могут существовать неизвестные патогены, которые могут прямо или косвенно играть роль в энтерите у индеек. Этот обзор будет посвящен роли коронавируса, ротавируса, реовируса и астровируса индейки в энтерите индейки.
Ключевые слова: Энтерит, Индейки, Коронавирус, Ротавирус, Реовирус, Астровирус
Введение
На протяжении многих лет у индеек было описано несколько кишечных заболеваний, а именно: коронавирусный энтерит индеек, синдром мальдигестии, синдром ранения и задержки роста у индеек, синдром мальабсорбции у домашней птицы, синдром энтерита и смертности домашних птиц (PEMS), пиковая смертность индеек и вирусный энтерит индейки.Все эти синдромы были включены в комплекс энтерита домашней птицы (PEC), общий термин, обозначающий инфекционные кишечные заболевания молодняка индеек [6]. ПЭК — это экономически важное заболевание молодняка индеек, характеризующееся энтеритом, диареей, умеренной или выраженной депрессией роста, нарушением использования корма, плохой прибавкой веса и в некоторых случаях высокой смертностью, как при ПЕМС [6]. Ряд вирусов (коронавирус, калицивирус, реовирус, астровирус, ротавирус, пикорнавирус, пикобирнавирус, парвовирус и аденовирус), бактерий ( Escherichia coli , Salmonella spp., Clostridia, Campylobacter и Enterococcus ) и простейшие (кокцидии и криптоспоридиумы) участвуют в PEC [6, 18, 47, 73, 105, 109]. Недавно у индюшат Миннесоты были описаны два новых синдрома, а именно «синдром энтерита домашней птицы», или PES, и «синдром легкой индейки», или LTS. ПЭС — это инфекционное кишечное заболевание молодых индеек в возрасте от 1 дня до 7 недель, которое характеризуется диареей, депрессией и летаргией с бледным кишечником и / или чрезмерно жидким содержимым слепой кишки [47].LTS — это проблема индеек товарного возраста, имеющих меньшую массу тела по сравнению с их стандартными породными характеристиками [68]. Мы полагаем, что энтерит у домашних птиц в молодом возрасте создает условия для будущего развития LTS у более старых птиц. Кроме того, мы рассматриваем как PES, так и LTS как часть PEC.
Кишечные вирусы являются частой причиной первичного повреждения желудочно-кишечного тракта (ЖКТ) молодых цыплят, создавая благоприятную среду для роста бактерий и / или простейших и вызывая дальнейшее повреждение кишечника.Возможно, что повреждение кишечника сначала кишечными вирусами, а затем вторичными патогенами может привести к необратимым изменениям в организме хозяина, что может привести к снижению массы тела при продаже. Цель этого обзора — подробно обсудить четыре наиболее важных вируса, вызывающих энтерит индейки, а именно коронавирус, реовирус, ротавирус и астровирус.
Коронавирус
Коронавирусный энтерит индейки — острое заразное заболевание, характеризующееся тупостью, депрессией, диареей и сниженным набором массы тела.Заболевание было впервые описано в 1940-х годах в штате Вашингтон, США. Возбудитель не был идентифицирован, и заболевание было названо «грязевой лихорадкой». Однако после выявления в этих случаях коронавируса индейки (TCoV) название было изменено на «коронавирусный энтерит индеек». В 1970-х годах эта болезнь имела самые разрушительные последствия для индеек из Миннесоты. Были предприняты шаги по искоренению болезни, которые привели к элиминации этого вируса из Миннесоты. В настоящее время TCoV спорадически регистрируется из регионов Северной Америки и других стран, где выращивают индейку.TCoV также связан с PEMS, острым высококонтагиозным кишечным заболеванием, характеризующимся депрессией, анорексией, диареей и высокой смертностью индеек.
Геном
Коронавирусы представляют собой плеоморфные частицы с оболочкой, примерно сферической формы, диаметром от 50 до 200 нм. Согласно последнему предложению Международного комитета по таксономии вирусов (ICTV), семейство Coronaviridae включает два подсемейства: Coronavirinae и Torovirinae. Coronavirinae состоит из трех родов: Alphacoronavirus , Betacoronavirus и Gammacoronavirus [54], из которых последний включает коронавирусы птиц. Геном коронавируса состоит из одноцепочечной РНК с положительным смыслом. Полноразмерный геном состоит из 27 632 нуклеотидов плюс 3 ‘поли (А) хвост. Есть две открытые рамки считывания (ORF), ORF 1a и 1b, которые находятся в первых двух третях генома с девятью дополнительными ORF ниже по течению.Область между генами белков мембраны (M) и нуклеокапсида (N) содержит три потенциальных малых ORF: (1) ORF-X, (2) ранее не охарактеризованную ORF со связанной предполагаемой TRS в гене M (очевидно, общей для всей группы III coronaviruses) и (3) ранее описанные ORF 5a и 5b. TCoV не содержит гена гемагглютинин-эстеразы [29].
Сообщалось о вариабельности в различных генных сегментах TCoV. Сравнение последовательностей полной 3′-концевой области, кодирующей структурный белок четырех TCoV, с соответствующими последовательностями вируса инфекционного бронхита (IBV) показало, что TCoV, вероятно, имеет то же происхождение, что и IBV, и приобретенные последовательности S-гена для тропизма кишечника индейки во время процесс эволюции [61].Lin et al. [60] сообщили, что последовательность S-белка TCoV имеет только 33,8–33,9% идентичности с IBV, хотя оба вируса сгруппированы в одну и ту же геномную линию. Полноразмерные последовательности гена spike (S) французского происхождения TCoV обладали 98% идентичностью как на уровне нуклеотидов, так и на уровне аминокислот [65]. Однако эти вирусы показали 65 и 60% сходства с североамериканскими TCoV и 50 и 37% сходства с IBV на уровне нуклеотидов и аминокислот, соответственно.
Заболеваемость и распространение
В отличие от обычно неопределенных синдромов вирусных кишечных заболеваний, TCoV-энтерит можно напрямую отнести к инфекции, вызванной TCoV [35, 38, 41].TCoV распространен во всем мире и был обнаружен у пораженных энтеритом индеек из Северной Америки, Южной Америки и Европы [34, 59, 65, 70, 98]. Было обнаружено, что 37% кишечных проб от больных стад индейки в Европе оказались положительными на TCoV [65]. В недавнем исследовании, проведенном в Бразилии, большинство образцов кишечника, взятых как у пораженных энтеритом, так и у здоровых индеек, оказались положительными на TCoV [70]. В этом исследовании 55,3% TCoV-положительных образцов также были положительными на другие кишечные вирусы.Недавно были зарегистрированы вспышки коронавирусного энтерита из Арканзаса и Северной Каролины в США с тяжелой диареей и смертностью [19, 83, 100, 106], что указывает на важность постоянного эпидемиологического надзора за этим вирусом. TCoV, по-видимому, отсутствует в синдромах ПЭК без ПЭМС [44, 45, 62, 68, 75, 105]. Например, коронавирус не был обнаружен, когда в Калифорнии были протестированы 2400 образцов от домашних птиц с клиническими признаками энтерита [105]. Аналогичным образом, в исследовании 33 стай индеек, пораженных энтеритом, в США, Pantin-Jackwood et al.[75] не нашли TCoV. Мы также не обнаружили TCoV в случаях PES и LTS [44, 45, 68].
TCoV может поражать птиц любой возрастной группы, но вызывает высокую смертность молодых индюшат в возрасте 1–4 недель. Смертность колеблется от менее 10 до 50%. Индюки, по-видимому, являются единственным естественным хозяином TCoV; при экспериментальной инокуляции TCoV вызывал заболевание у домашних птиц, но не у цыплят [42]. Репликация TCoV происходит в энтероцитах, выстилающих апикальную часть ворсинок кишечника. Разрушение эпителия ворсинок кишечника приводит к диарее, нарушению пищеварения и мальабсорбции, что может привести к снижению прибавки в весе у индюшат.
Передача
Вирус присутствует в основном в кишечнике и фекалиях пораженных домашних птиц, следовательно, передача вируса осуществляется фекальным оральным путем. Прямая передача происходит при прямом контакте с инфицированными птицами, тогда как косвенная передача происходит механическим путем через рабочих и / или зараженное оборудование. Однако в экспериментальном исследовании антиген TCoV также может быть обнаружен в придаточных пазухах носа и слезной добавочной железе (железе Хардера) инфицированных цыплят в течение 14 дней после инокуляции [33].Вирус выделяется с фекалиями индеек в течение многих недель после выздоровления от клинического заболевания. Большинство случаев заражения происходит летом (май – август), иногда осенью. TCoV может сохраняться в течение длительного периода времени и может легко передаваться между птицефабриками в прохладной окружающей среде.
Клинические признаки и патология
Индюки, пораженные TCoV, сбиваются в кучу и демонстрируют взъерошенные перья, снижение потребления корма и воды, влажный помет и потерю живой массы.У пожилых птиц может наблюдаться депрессия, диарея и задержка роста [34]. Помет водянистый и пенистый, может содержать слизь, от зеленого до коричневого цвета. Клинические признаки могут сохраняться до 2 недель, а восстановление веса, если оно будет достигнуто, может занять несколько недель. У несушек наблюдается снижение яйценоскости [3] из-за аномальной пигментации яиц. Птицы в возрасте от 1 до 4 недель, у которых наблюдаются диарея, обезвоживание и задержка роста на 40% или более, считаются страдающими PEMS. Существует две формы PEMS: (1) тяжелая форма i.е. рост нравственности индеек (SMT), который определяется как смертность, равная или превышающая 1% в течение 3 дней подряд или 9% в возрасте от 8 до 28 дней; и (2) легкая форма, которая определяется как смертность более 2%, но менее 9% в течение того же периода времени, но не равная или не превышающая 1% в течение трех дней подряд. В экспериментальном исследовании, как заболеваемость, так и смертность регистрировались у домашних птиц, экспериментально инокулированных TCoV в возрасте 2 дней, однако птицы, инфицированные в возрасте 28 дней, имели значительную задержку роста, но не демонстрировали смертности [32].
TCoV может действовать синергетически с другими патогенами, увеличивая тяжесть заболевания, или может предрасполагать птиц к вторичным инфекциям. У цыплят, инокулированных только TCoV, развилась умеренная депрессия роста без гибели, а у свиней, инокулированных только энтеропатогенной Escherichia coli (EPEC) , клиническое заболевание не развилось. Однако инокуляция цыплят TCoV и EPEC одновременно вызвала повышение смертности, подавление роста и прикрепление / стирание повреждений [38].
У пораженных птиц наблюдается задержка роста и обезвоживание. Кишечник заметно увеличен и заполнен рыхлым желтым содержимым. Стенки кишечника вялые и бледные. Caeca также вздуты с пенистым и водянистым содержимым. Крупные изменения, такие как атрофия поджелудочной железы, селезенки и бурсы Фабрициуса, также могут наблюдаться наряду с расширением желчного пузыря и мочеточников. У инфицированных кур наблюдаются более выраженные поражения яичников с деформированными, плохо развитыми яичниками и скоплением казеозного экссудата в брюшине.Также наблюдаются спайки брюшины яйцевода.
В зависимости от тяжести инфекции наблюдается от легкого мультифокального до тяжелого энтерита с атрофией ворсинок. Наблюдается слущивание эпителиальных клеток с застоем и обширной инфильтрацией лимфоцитов и гетерофилов в собственную пластинку ворсинок. Кроме того, также может наблюдаться истощение фолликулярных лимфоидов и атрофия коркового вещества тимуса.
Диагноз
Клинические признаки, такие как диарея, депрессия, скученность и задержка роста, указывают на кишечную инфекцию.Высокая смертность молодняка может указывать на ПЕМС.
Выделение вируса, электронная микроскопия (ЭМ), обнаружение вирусной РНК и серология могут помочь в диагностике. Вирус можно выращивать в зародышах индейки или куриных яйцах, но его трудно выращивать в клеточных культурах. ЭМ с отрицательным контрастом может выявить вирусные частицы размером ~ 50–200 нм с типичной морфологией в форме короны в содержимом кишечника. Прямые и непрямые флуоресцентные тесты на антитела и ELISA использовались для обнаружения антигена TCoV и антител соответственно [30, 31, 37].О высокой распространенности TCoV сообщалось от заводчиков (71,1%) и мясных индеек (56,7%) в Онтарио, Канада [31]. В Арканзасе 64,2% образцов сыворотки дали положительный результат на антитела [30].
Доступны молекулярные тесты для обнаружения и дифференциации TCoV от других кишечных вирусов. Доступны различные тесты полимеразной цепной реакции с обратной транскрипцией (RT-PCR), нацеленные на 3′-нетранслируемую область (UTR), S, N и / или M ген TCoV [11, 12, 16, 89, 92, 104]. Также была разработана ОТ-ПЦР в реальном времени, которая является наиболее чувствительным и специфическим тестом, с помощью которого можно количественно определить вирусную нагрузку в тканях индейки, а также в образцах фекалий [15].Прямая иммуногистохимия была рекомендована в качестве альтернативы ОТ-ПЦР для обнаружения TCoV в полевых образцах [14].
Профилактика и контроль
Специального лечения или вакцины нет. Биобезопасность, при которой особое внимание уделяется управлению подстилкой и надлежащей утилизации мертвых птиц и использованного подстилки, похоже, единственный способ предотвратить распространение инфекции. Кроме того, общие меры, такие как уменьшение влажности подстилки, использование антибиотиков для борьбы с вторичными бактериальными инфекциями и улучшение гигиены, могут минимизировать последствия энтерита TCoV.
Реовирус
Птичий реовирус был впервые выделен от кур, страдающих хроническим респираторным заболеванием, и получил название возбудитель Фейи-Кроули (FC). Позже возбудитель FC был охарактеризован как реовирус. Однако у кур птичий реовирус вызывает множество синдромов, включая вирусный артрит / теносиновит. Недавно реовирусы были выделены от артрита / тендосиновита индейки и получили название TARV (реовирус артрита индейки) [69]. Реовирусы в основном бывают двух типов: фузогенные и нефузогенные.Слитые вирусы обладают способностью вызывать слияние инфицированных клеток, что приводит к образованию многоядерных синцитий, в то время как нефузогенные вирусы не вызывают слияния инфицированных клеток. К нефузогенным типам относятся, в основном, реовирусы млекопитающих (MRV), тогда как слитные реовирусы поражают млекопитающих, птиц и рептилий.
Геном
Реовирус принадлежит к роду Orthoreovirus в семействе Orthoreoviridae. Вирус без оболочки, имеет икосаэдрическую симметрию и размер частиц 70–80 нм.Он содержит геном двухцепочечной (ds) РНК из десяти сегментов, которые делятся на три класса, называемые большими (L), средними (M) и малыми (S), в зависимости от их характера миграции при электрофорезе в полиакриламидном геле. Гены L и M далее подразделяются на три сегмента каждый (L1, L2, L3 и M1, M2, M3, соответственно), в то время как ген S имеет четыре сегмента (S1, S2, S3, S4) [7].
Геном реовируса имеет 12 открытых рамок считывания, которые кодируют восемь структурных и четыре неструктурных белка.Структурные белки являются важной частью вирионов потомства, в то время как неструктурные белки не присутствуют в зрелом вирионе и экспрессируются только в инфицированных клетках. Структурными белками, кодируемыми генами L, M и S, являются лямбда (λ), мю (μ) и сигма (σ) соответственно. Три структурных белка λA, λB и λC кодируются сегментами L гена L1, L2 и L3 соответственно. Сегменты M1 и M2 кодируют два структурных белка (μA и μB) соответственно, в то время как сегмент M3 кодирует неструктурный белок (μNS).Три σ-белка σC, σA и σB кодируются сегментами S1, S2 и S3 соответственно, тогда как сегмент S4 кодирует неструктурный белок σNS. Сегмент S1 кодирует два дополнительных неструктурных белка, а именно p10 и p17 [7, 10].
На основании анализа последовательностей различных генов сообщалось, что реовирусы индейки отличаются от реовирусов кур [20, 44, 45, 52, 88]. Например, сходство нуклеотидной и аминокислотной последовательности сегмента S3, кодирующего сигма-B-белок реовируса индейки (TRV), было обнаружено примерно 61 и 78-80%, соответственно, по сравнению с изолятами куриного реовируса [88].Day et al. [20] сообщили, что белок сигма C гена S1 TRV имеет 57% аминокислотную идентичность с таковой у куриного реовируса (эталонный штамм S1133), тогда как белки p10 и p17 имеют 72 и 61% идентичности, соответственно, с соответствующими белками S1133.
Белок сигма С реовируса млекопитающих и слитый реовирус Нельсона Бэй на 25–28% имеют аминокислотную идентичность с белком сигма С TRV. Ранее мы сообщали, что сходство последовательностей гена S4 на уровне нуклеотидов TRVs было 71.1–74,2% по сравнению с реовирусами куриного происхождения [45]. Эти исследования показывают, что TRV образуют отдельную группу по сравнению с куриными и другими изолятами птичьего реовируса. В результате TRV потенциально можно рассматривать как отдельный вид вируса в подгруппе 2 рода Orthoreovirus . Эта точка зрения дополнительно подтверждается экспериментальными исследованиями, в которых TRV не смогли вызвать признаки заболевания у цыплят, хотя они могут реплицироваться на низких уровнях в кишечном тракте цыплят.Более того, высокопатогенный штамм реовируса куриного происхождения (ARV-1733), вызывающий высокую смертность цыплят, не вызывал клинических признаков у промышленных индеек и имел лишь легкие клинические признаки у индюков, свободных от специфических патогенов [93].
Заболеваемость и распространение
Около 85–90% изолятов реовируса не являются патогенными. Патогенные штаммы реовирусов были связаны с различными заболеваниями кур и индеек. У индеек этот вирус был обнаружен при нескольких различных кишечных заболеваниях, а именно LTS, PEC, PES, миокардите, а недавно и при артрите / теносиновите [6, 28, 39, 44, 47, 68, 69, 90, 91].При исследовании 33 стад индюшат 46% стад оказались положительными на реовирусы [75]. Jindal et al. [44] проверили содержимое кишечника 43 стад, пораженных PES, в 2007–2008 гг. И сообщили о наличии реовируса в 40% стад. Энтерит, вызванный реовирусом, в основном встречается у молодняка индейки в возрасте 1–7 недель, а заболеваемость выше у птиц в возрасте 1–3 недель, которая уменьшается с увеличением возраста.
Реовирусы обнаружены также у практически здоровых кур и индеек [45, 68, 75].При исследовании пяти здоровых родительских стад индюшат 10,4% из 193 протестированных образцов (в возрасте от 1 до 9 недель) были положительными на реовирус [45]. Тестирование образцов кишечника 33 промышленных стад индейки из всех регионов США в течение 2005 и 2006 гг. Выявило присутствие реовируса в 45,5% стад [75].
Передача
Горизонтальная передача фекально-оральным путем является обычным явлением. Из-за стабильности вируса в окружающей среде также может происходить механическая передача.Различные штаммы реовируса различаются по способности распространяться по горизонтали. Подозревается, но не доказано, вертикальная передача кишечной реовирусной инфекции.
Клинические признаки и патология
Пораженные птицы страдают диареей, депрессией, взъерошенными перьями, а также сниженным набором веса [93] и смертностью. Инфекция АРВ может также предрасполагать птиц к вторичным осложнениям или действовать синергетически вместе с другими инфекциями. Сообщалось, что АРВ-инфекция может усиливать кокцидиальную инфекцию у цыплят-бройлеров [85].Также существует возможность синергического действия TRV с другими патогенами у индеек.
В большинстве случаев содержимое кишечника пенистое и водянистое. В некоторых случаях наблюдается атрофия бурсы Фабрициуса [22, 93]. Поражения кишечника состоят из легкой гиперплазии крипт из-за инфильтрации лимфоцитов, гетерофилов и эозинофилов в собственную пластинку и подслизистую оболочку через 11–14 дней после заражения. В сумке Фабрициуса наблюдается умеренное или тяжелое истощение фолликулярных лимфоцитов [22, 93].Помимо атрофии бурсальной сумки, может наблюдаться лимфоидное истощение от легкой до умеренной степени в селезенке и лимфоцитарная инфильтрация в печени, поджелудочной железе, сердце и преджелудках.
Диагноз
Диарея в стаде является признаком кишечной инфекции. Трудно точно определить вирус / возбудитель только на основе диареи, и для подтверждения необходимы лабораторные тесты. Электронная микроскопия может выявить присутствие вирусных частиц размером ~ 70–80 нм в кишечнике инфицированных птиц.Вирус хорошо растет в курином яйце с эмбрионом через желточный мешок или хориоаллантоисную мембрану, а также был выделен в первичных клетках печени куриного эмбриона (CEL), клетках почек куриного эмбриона (CEK) и клетках фибросаркомы японского перепела (QT-35). Из-за фузогенной природы TRV в инокулированных культурах клеток наблюдается характерный цитопатический эффект (ЦПЭ). Для обнаружения антител против реовируса цыплят коммерчески доступны наборы ELISA, но в настоящее время таких специфических наборов для реовируса индейки нет.Недавно мы разработали тест ELISA для обнаружения антител против реовируса индейки. В этом тесте мы клонировали и экспрессировали белок сигма C S1 и использовали его в качестве антигена для обнаружения антител против TRV (неопубликованные данные Mor et al.).
Доступно несколько тестов ОТ-ПЦР, нацеленных на различные генные сегменты реовируса [4, 20, 44, 45, 52, 88]. ОТ-ПЦР в реальном времени — это наиболее чувствительный и специфический тест, с помощью которого можно количественно определить вирусную нагрузку в клинических образцах. Был разработан надежный, сверхчувствительный и точный количественный анализ на АРВ с помощью Light Cycler SYBR Green на основе ОТ-ПЦР в реальном времени, который может обнаруживать 39 копий / мкл геномной РНК АРВ [53].Недавно в нашей лаборатории была разработана высокочувствительная и специфическая ОТ-ПЦР в реальном времени для обнаружения TRV. Мы разработали праймеры и зонды из сегмента генома S4. Тест специфичен для TRV (энтерических вирусов индейки и реовирусов артрита) и может обнаруживать до 10 копий транскрибированной in vitro стандартной вирусной РНК на реакцию (Mor et al. Неопубликованные данные).
Профилактика и контроль
Эффективного лечения реовирусных инфекций не существует. Вирус является повсеместным по своей природе, и из-за его высокой стабильности в окружающей среде трудно избавить птиц и фермы от реовируса.В настоящее время вакцины против реовируса индейки нет, и поэтому биозащита остается единственным способом борьбы с инфекцией.
Ротавирус
Ротавирусы являются важными патогенами, которые вызывают неонатальную диарею у людей, а также у многих животных и птиц. Этот вирус также был обнаружен у коммерческих цыплят-бройлеров, страдающих синдромом короткого роста и задержки роста [72].
Геном
Ротавирусы — это РНК-вирусы, принадлежащие к роду Rotavirus и семейству Reoviridae .Геном ротавируса состоит из 11 сегментов двухцепочечной РНК; РНК окружена тройным слоем икосаэдрического белкового капсида. Каждый сегмент кодирует как минимум один белок. Внешний белковый слой состоит из двух основных нейтрализующих антигенов — вирусного белка 4 (VP4) и вирусного белка 7 (VP7). Вирусный белок 6 (VP6) второго слоя капсида называется групповым антигеном, который используется для обнаружения и классификации ротавирусов на отдельные группы. Кроме того, по крайней мере пять неструктурных белков кодируются геномом ротавируса.
Основываясь на антигенных свойствах VP6, ICTV разделил ротавирусы на пять серологических видов (A – E) и два дополнительных вида (F и G) [81], которые также называются группами ротавирусов. Известно, что группы A, B и C инфицируют людей и животных, а группы D, E, F и G заражают только животных, в основном птиц [50, 63, 102]. Недавно было сообщено о потенциально новом виде RV, ротавирусе H [ 64 ]. Эти авторы утверждают, что значение отсечения аминокислотной последовательности 53% позволяет дифференцировать различные виды ротавирусов.
В отличие от ротавирусов птиц группы A, ротавирусы группы D не могут размножаться в клетках MA104 [ 25 ] и в основном идентифицируются по схеме электрофоретической миграции 5: 2: 2: 2 их сегментов генома, что значительно отличается от паттерна 5: 1: 3: 2 ротавирусов птичьей группы А. Однако классификация штамма ротавируса только на основе его профиля миграции РНК является проблематичной из-за возможного возникновения перестройки генома. В отличие от ротавирусов группы А, которые преимущественно инфицируют клетки двенадцатиперстной кишки, вирусы группы D имеют склонность к тощей и подвздошной кишке.
Ротавирус индейского происхождения отличается от ротавируса куриного происхождения [58]. Например, сообщалось, что гомология аминокислотной последовательности гена VP7 штамма Ty-1 (индейка) и гена Ch-2 VP7 птиц составляет только 70% [58]. Вариабельные эпитопные области A, B и C Ty-1 были уникальными по сравнению с таковыми у Ch-2 и других штаммов, представляющих серотипы 14G. Низкая гомология (53%) областей A и C Ty-1 и Ch-2 предполагает, что штамм Ty-1 может иметь другой серотип, чем эталонный штамм G7 Ch-2.Сравнение последовательностей различных генов (VP4, VP6, VP7, NSP5) ротавирусов индейки и кур показало, что среди ротавирусов птичьей группы А может происходить межвидовая передача и реассортация [87].
Заболеваемость и распространение
Ротавирусы распространены по всему миру и были зарегистрированы как у здоровых, так и у пораженных энтеритом индеек [44, 45, 69, 71, 75, 77, 99]. Периодический мониторинг товарных стад индейки выявил циркуляцию четырех групп (на основе анализа последовательности гена NSP4) ротавирусов.Среди четырех групп ротавирусов было 96,1–97,5 и 97,5–99,55% сходства на уровне нуклеотидов и аминокислот соответственно [77]. Ротавирусы присутствовали до размещения, а также после размещения до 12-недельного возраста [77]. В другом исследовании, в котором участвовали 33 стада индейки и 43 стада кур, ротавирусы были обнаружены в 46% стадах цыплят и 70% стадах индейки [75]. Мы также сообщали о присутствии ротавирусов в практически здоровых стадах производителей индейки с максимальной встречаемостью до 5-недельного возраста [45].В другом исследовании 93% стад, пораженных PES, были положительными на ротавирусы [44]. В контрольном исследовании Mor et al. [68] сообщили, что шесть (7,5%) пулов образцов фекалий были положительными на ротавирус в стадах, пораженных LTS, в то время как 13 (33%) пулов образцов были положительными из стада без LTS.
Выявление ротавирусов выше у молодых цыплят, и уровень обнаружения снижается с возрастом. При наблюдении PES и LTS максимальное количество цыплят было положительным на ротавирус в возрасте 2–5 недель с последующим уменьшением количества положительных образцов в возрасте 8–9 недель [44, 45, 68].Было показано, что праймеры на основе высококонсервативной области генома ротавируса (в частности, неструктурных белков NSP3 или NSP4) подходят для идентификации ротавируса в различных клинических образцах, образцах животных или окружающей среды с помощью обычных или в реальном времени RT- ПЦР [51, 84]. В исследовании, проведенном в Европе, ротавирусы кур и индеек были идентифицированы с использованием двух разных методов: электрофореза в полиакриламидном геле с ОТ-ПЦР и анализов ОТ-ПЦР в реальном времени, специфичных для ротавирусов групп A и D [71].Эти результаты показывают, что необходимы дальнейшие исследования для точного определения типов ротавирусов, циркулирующих в стадах индейки в различных географических точках.
Передача
Фекальный оральный путь является основным путем горизонтальной передачи. Вирус может выжить в окружающей среде в течение длительного времени, и, следовательно, механическая передача может играть роль в его передаче. Есть подозрение на вертикальную передачу ротавируса, но это пока не подтверждено. Например, Pantin-Jackwood et al.[77] обнаружили ротавирусы в 50% проб, исследованных до помещения птицы. Основываясь на этой информации, авторы предположили, что ротавирус может передаваться вертикально. Мы также обнаружили ротавирусы у 2-дневных коммерческих и родительских домашних птиц [45, 47], что указывает на возможность вертикальной передачи ротавируса.
Также описана межвидовая передача ротавируса [87, 103]. В одном исследовании ротавирусы крупного рогатого скота группы А (n = 6) были обнаружены в кишечном содержимом индюков [2].Электроферотипы показали картину миграции, идентичную вирусу диареи телят в Небраске, а полная филогения гена NSP4 показала, что все шесть штаммов сегрегированы по генотипу E2, что подтверждает их принадлежность к ротавирусу крупного рогатого скота. Ротавирусы птиц не обнаружены у людей, и ротавирусы человека не содержат последовательностей ротавирусов птиц [87], что указывает на низкую вероятность заражения человека ротавирусами птиц.
Клинические признаки и патология
Тупость, депрессия, водянистый помет, потеря аппетита, склеивание вентиляционных отверстий и снижение прибавки в весе — постоянные признаки у пораженных птиц.Смертность может присутствовать или отсутствовать, а заболеваемость может варьироваться. Ротавирусная инфекция кур-несушек может привести к снижению яйценоскости.
В большинстве случаев наблюдается пенистое и водянистое кишечное содержимое. Может наблюдаться бледность кишечного тракта и вздутие слепой кишки с пенистым или непенистым жидким содержимым. Другими крупными патологическими проявлениями являются обезвоживание, задержка роста и пастообразные выделения. Также отмечаются мультифокальные и поверхностные эрозии двенадцатиперстной и тощей кишки. Наиболее частыми микроскопическими изменениями являются гиперцеллюлярность собственной пластинки из-за инфильтрации гетерофилов, эозинофилов и мононуклеарных клеток, а также атрофия ворсинок.Сканирующая электронная микроскопия показывает шероховатость поверхности ворсинок, нарушение нормальной морфологии ворсинок и потерю микроворсинок в клетках, расположенных на кончиках ворсинок. У цыплят разные штаммы ротавирусов могут иметь разные предпочтительные сайты репликации. Например, McNulty et al. [66] сообщили, что у экспериментально инфицированных кур ротавирус группы A имел сродство к двенадцатиперстной кишке, тогда как ротавирус группы D предпочитал тощую кишку и подвздошную кишку.
Диагноз
Диарея, обезвоживание и снижение веса стада свидетельствуют о кишечной инфекции.ЭМ кишечного содержимого и фекалий выявляет присутствие вирусных частиц размером ~ 70 нм (приблизительно 50–70 нм) с характерным «колесом» внешнего вида. Несмотря на то, что вирус трудно изолировать, он все же растет в клетках MA104 при обработке трипсином. Тест коагглютинации использовался для обнаружения ротавирусов у индеек и оказался простым и быстрым скрининговым тестом. Электрофорез в полиакриламидном геле обычно используется для определения характерной картины миграции 11 сегментов генома ротавируса и почти так же чувствителен, как ЕМ.
Для обнаружения вируса доступно несколько ОТ-ПЦР и ОТ-ПЦР в реальном времени, нацеленных на различные сегменты генома ротавируса [8, 21, 77]. Недавно было сообщено о высокочувствительной и специфической одностадийной ОТ-ПЦР в реальном времени, нацеленной на неструктурный белок 4 (NSP4), с внутренней системой контроля для обнаружения ротавирусов индейки [1].
Профилактика и борьба
Специальных мер борьбы с ротавирусными инфекциями не существует. Получение цыплят из источников, свободных от ротавирусов, и принятие надлежащих мер биобезопасности — это способы минимизировать заражение ротавирусом в стаде.
Астровирус
Астровирус индейки (TAstV) был впервые обнаружен у индюшат в возрасте 6–11 дней с диареей и повышенной смертностью в Великобритании. В США TAstV был обнаружен у индюшат и позже был назван TAstV-1. Впоследствии TAstV, связанный с PEMS, был выделен и охарактеризован. Этот вирус генетически и иммунологически отличался от ранее описанного изолята США, TAstV-1, и был назван TAstV-2 [56, 86]. Помимо TAstV-1 и TAstV-2, астровирус куриного происхождения i.е. вирус птичьего нефрита (ВНВ) обнаружен также у индеек [26, 75]. ANV сначала был описан как небольшой круглый вирус, затем как пикорнавирус, а позже был определен как астровирус [40]. Таким образом, у индеек обнаружено три типа астровирусов (TAstV-1, TAstV-2 и ANV). Однако данные о последовательностях предполагают, что в природе может присутствовать гораздо больше типов [94]. В 1980-х годах все обнаруженные астровирусы были строго ограничены желудочно-кишечным трактом домашних птиц, и не было никаких признаков системного заболевания.Однако TAstV также был обнаружен в тимусе и сумке, что указывает на возможное влияние на иммунную систему [86]. Сообщалось также об обнаружении астровирусов у кур и индюшат, страдающих кишечными и / или опорно-двигательными расстройствами (теносиновит) [26, 82].
Геном
Астровирусы — это РНК-вирусы без оболочки диаметром 25–35 нм, принадлежащие к роду Avastrovirus семейства Astroviridae . Геном астровируса состоит из 6,8–7 РНК.9 kb и имеет 5′-UTR, за которым следуют три ORF, 3′-UTR и поли-A-хвост. ORF-1a кодирует сериновую протеазу и другие неструктурные белки, ORF-1b для РНК-зависимой РНК-полимеразы и ORF-2 для капсидного белка. Общая длина геномов трех астровирусов, а именно. TAstV-1 (7003 нт), TAstV-2 (7325 нт) и ANV (6927 нт) варьируются; эта длина не включает хвост поли-А [55].
Большинство исследований по обнаружению и характеристике TAstV было проведено молекулярными методами с использованием праймеров, специфичных для гена капсида или полимеразы.Ряд исследований показал вариации на уровне нуклеотидов и аминокислот в гене капсида и / или гене полимеразы TAstVs [13, 44–46, 78, 79, 97]. Сообщается о высоком уровне генетической изменчивости среди астровирусов индейки из стад индейки в США [78]. Идентичность нуклеотидной последовательности в гене капсида, связанная с серотипом и вирусным патогенезом, была всего лишь 69%, но ген полимеразы был более консервативным с 86–99% идентичностью нуклеотидов. Недавно филогенетический анализ белка-предшественника капсида выявил обширные вариации в аминокислотных последовательностях TAstV-2 (81.5–100%), но не в TAstV-1 (96,2–100%) [79]. Мы также обнаружили вариации на уровне аминокислот в гене капсида TAstV-2 [44, 46]. Эти изменения предполагают, что в природе могут существовать различные серотипы астровируса индейки. Для лучшего понимания серотипов TAstV, циркулирующих у индеек, необходимы исследования полных последовательностей генов капсида из явно здоровых и пораженных PEC стад из разных географических регионов. Изменения гена капсида из-за мутаций или рекомбинации могут влиять на антигенность и патогенность вирусов и, следовательно, иметь практическое значение для методов обнаружения вирусов, эпидемиологических исследований и разработки потенциальных вакцин против астровирусных инфекций.
Заболеваемость и распространение
Был проведен ряд исследований для выявления присутствия TAstV в стадах индейки с использованием различных методов. Ранее вирусы в образцах кишечника выявлялись с помощью прямой или иммунной ЭМ. Однако в большинстве недавних исследований используются молекулярные инструменты, такие как ОТ-ПЦР и секвенирование, для обнаружения различных астровирусов [36, 56, 57, 108]. В ходе исследования восьми промышленных предприятий по выращиванию индейки и одного исследовательского подразделения в США, 89,5% из 96 образцов содержимого кишечника были положительными на астровирус с помощью ОТ-ПЦР [77].В исследовании, проведенном в Миннесоте, 84% из 43 стай индеек, пораженных PES, оказались положительными на TAstV-2 с помощью ОТ-ПЦР [44]. Эти образцы не тестировались на наличие TAstV-1 и ANV, поэтому нельзя исключать присутствие TAstV-1 и ANV в этих образцах. Ряд других исследований также выявил высокую распространенность TAstV в стадах индейки в США [75, 77, 78].
Исследования распространенности TAstV также проводились в других странах, где выращивают индюшат [17, 27, 62, 70, 74]. Да Силва и др.[17] сообщили о TAsV у молодых цыплят, пораженных PEC в Бразилии. В другом исследовании, проведенном в Бразилии, TAsV-1, TAstV-2 и ANV были идентифицированы в 64,5, 44,7 и 35,5% образцов кишечника соответственно [70]. Из них TAstV-1 и TAstV-2 были более распространены в стадах индеек, пораженных энтеритом. Domanska-Blicharz et al. [27] сообщили, что TAstV-2 является наиболее распространенным вирусом в Польше; 30 из 77 стад индейки (38,9%) были положительными. TAstV-1 был обнаружен только в девяти стадах, тогда как ANV — в одном стаде. Эти три вируса были обнаружены либо как одиночные, либо как смешанные инфекции.Тестирование 23 образцов кишечника от откормленных индюков с клиническими признаками энтерита выявило присутствие TAstV-2 в 17 образцах в Хорватии [62].
TAstV были обнаружены не только в стадах индюшат, пораженных энтеритом, но также и в практически здоровых стадах индюшат [45, 68, 78]. Jindal et al. [45] протестировали 193 объединенных образца фекалий практически здоровых индюшат-производителей с помощью ОТ-ПЦР и обнаружили, что 47,2% оказались положительными на TAstV-2.
Передача
Передача астровирусов фекально-оральным путем.Здоровые птицы могут подвергаться воздействию астровируса либо при прямом контакте с инфицированными фекалиями, либо с материалами, загрязненными фекалиями. Считается, что астровирусные инфекции видоспецифичны, но недавние сообщения указывают на межвидовую передачу [23, 101]. Так, TAstV-2 и ANV были обнаружены у уток ( Anas platyrhynchos domesticus ) [9]. Межвидовая передача и адаптация вируса к новым хозяевам или совместное инфицирование одного и того же хозяина разными астровирусами может привести к появлению новых астровирусов, которые инфицируют животных или обладают зоонозным потенциалом [24].
Клинические данные и патология
Как и при рота- и реовирусных инфекциях, астровирусы также вызывают диарею, обезвоживание и задержку роста. Сообщалось о подобных наблюдениях диареи и задержки роста у экспериментальных цыплят, инокулированных материалом, содержащим TAstV [48, 49, 76]. Были предприняты экспериментальные исследования, чтобы установить тип клинического заболевания, вызываемого TAstV, имеющими вариантные капсиды, или те, которые различались по источнику происхождения. Например, Pantin-Jackwood et al.[76] изучали патогенность трех различных вирусов TAstV-2 у индюков, свободных от специфических патогенов, и обнаружили, что вирусы TAstV-2 с вариантными капсидами вызывают подобное кишечное заболевание у молодых индеек. Mor et al. [67] обнаружили, что TAstV-2 от птиц PES был более патогенным, чем TAstV-2 от практически здоровых домашних птиц. Астровирус может потенциально нарушать иммунный ответ, тем самым повышая восприимчивость индеек к вторичным бактериальным инфекциям [80].
Обезвоживание, вздутие кишечника, наполненное водянистым содержимым и непереваренной пищей, и расширенная слепая кишка с пенистым содержимым — это результаты вскрытия.Может возникнуть атрофия бурсальной сумки от легкой до умеренной, а также снижение тонуса кишечника. Гистологическими находками являются умеренный некроз эпителия, инфильтраты собственной пластинки и легкая гиперплазия крипт.
Диагноз
Диарея, обезвоживание и снижение веса являются признаками кишечной инфекции. Электронная микроскопия с отрицательным контрастом выявляет вирусные частицы с пятью или шестью выступами на поверхности, напоминающими звезду. Звездообразная морфология не всегда сохраняется или четко различима.По этой причине вирусы AstV от EM можно спутать с пикорнавирусами, пикорнавирусами, энтеровирусами и энтероподобными вирусами. Во-вторых, домашние птицы могут демонстрировать клинические признаки и патологию на более поздней стадии астровирусной инфекции, но в кишечном содержимом может не быть обнаруживаемых астровирусных частиц. В такой ситуации, даже если заболевание возникло, результаты анализов могут быть отрицательными. Молекулярные методы были использованы для обнаружения и дальнейшей характеристики астровирусов. В большинстве анализов на основе ОТ-ПЦР используются праймеры, специфичные для полимеразы или гена капсида TAstV [56, 57, 96].
Смешанные вирусные инфекции
Хотя отдельные вирусы были обнаружены в стадах индейки, пораженных энтеритом, большинство исследований указывают на присутствие двух или более энтеральных вирусов [44, 45, 62, 68, 70, 75, 77, 105, 107 ]. Периодический мониторинг восьми промышленных стад индейки в США выявил постоянное присутствие TAStV и ротавируса [77]. Из 96 протестированных образцов 89,5% были положительными на астровирус и 67,7% на ротавирус. Все стада были отрицательными на TCoV, реовирус и аденовирус группы 1 во всех точках отбора проб.В другом исследовании присутствие астровирусов было зарегистрировано в 100% протестированных стад индейки [75]. TAstV-2 и TAstV-1 были обнаружены в 100 и 15,4% стад индейки соответственно. Кроме того, 12,5% стад индейки были положительными на ANV. Ротавирусы и реовирусы присутствовали в 69,7 и 45,5% стад соответственно, при этом коронавирус и аденовирус не обнаружены.
Кишечное содержимое 43 стад PES в Миннесоте было проверено на наличие энтеральных вирусов методом RT-PCR.Все 43 стада оказались положительными на ротавирус, TAstV-2 или реовирус. Во всех случаях вирусы были обнаружены либо по отдельности, либо в комбинации из 2 или 3. Восемь стай (19%) были положительными на один вирус, а остальные 35 (81%) имели комбинацию вирусов. Пятнадцать стад оказались положительными на все три вируса. Ни одно из протестированных стад не было признано положительным на TCoV [44]. Исследования, проведенные в Бразилии, показали наличие 69,7% множественных вирусов в образцах кишечника индеек. Образцы кишечника чаще всего содержали 3–4 вируса на образец, причем вирусы из семейства астровирусов (TAstV-1, TAstV-2 или ANV, или их комбинация) присутствовали во всех образцах, которые дали положительный результат на вирусы [70 ].
Возможно, что комбинация вирусов может вызвать более серьезные побочные эффекты у домашних птиц, чем отдельные вирусы. В одном исследовании индюшатина, инокулированная комбинацией TAstV-2, реовируса индейки и ротавируса, имела значительно меньшую массу тела, чем контрольная группа [91]. В экспериментальных исследованиях мы также сообщали, что пероральная инокуляция цыплят материалом PES (положительным по ротавирусу, TAstV-2 и Salmonella) приводила к значительной задержке роста на срок до 50 дней после инокуляции, и ни в какой момент после инокуляции не наблюдалось. веса оспариваемых птиц сходятся с весом контрольных.Содержимое кишечника было водянистым и пенистым / пенистым (рис.,) У цыплят, инокулированных материалом PES [49]. Точно так же индюшатины, инокулированные содержимым кишечника птиц, пораженных LTS, имели значительно меньшую массу тела в возрасте 20 недель по сравнению с индюшатинами, инокулированными содержимым кишечника птиц без LTS [68]. Такая степень замедления роста из-за энтерита домашней птицы может привести к значительным экономическим потерям для производителей индейки, поскольку пораженная птица может не достичь ожидаемого веса при продаже.На уровне фермы нельзя исключать роль агентов, отличных от этих четырех распространенных вирусов (TAstV, ротавирус, реовирус и коронавирус), в возникновении и / или повышении серьезности кишечных вирусов. Кроме того, факторы питания и менеджмент также могут играть роль в возникновении энтерита у стада. Такие шаги, как улучшение методов лечения и противомикробная терапия для минимизации потерь, имели ограниченный успех [5]. Вакцины против кишечных вирусов в настоящее время нет.
Водянистое и пенистое содержимое слепой кишки птицы, инокулированной перорально кишечным материалом птиц, пораженных синдромом энтерита домашних птиц
Водянистое и пенистое содержимое слепой кишки птицы, инокулированной перорально кишечным материалом птиц, пораженных синдромом энтерита домашних птиц.Слепая кишка разрезана
Кишечные вирусы обычно обнаруживаются с помощью ЭМ, серологии и ПЦР. Большинство из этих вирусов трудно вырастить в зародышевых яйцах курицы / индейки и клеточных культурах [95]. Таким образом, было разработано несколько анализов одиночной и множественной RT-PCR (mRT-PCR) для обнаружения астровируса индейки, ротавируса индейки, коронавируса индейки и реовируса индейки [21, 43, 56, 77, 89, 92]. Мультиплексные анализы RT-PCR экономят время и деньги, поскольку два или более вируса могут быть обнаружены в одной реакции.
Общие выводы и направления на будущее
Кишечные вирусы (TAstV, ротавирус, реовирус и коронавирус) широко распространены в стадах индейки, но их клиническое значение неясно. Полное влияние этих вирусов на производительность стада требует тщательного изучения. Хотя кишечные заболевания, вызванные вирусами, являются экономическим бременем для фермеров, выращивающих индейки, ни один вирус не появился в качестве вероятного возбудителя, чтобы можно было сформулировать соответствующие меры по профилактике и контролю [18].В нескольких исследованиях одновременно были обнаружены два или более разных вируса, что указывает на то, что несколько вирусов могут участвовать в патогенезе энтерита. Преобладание вирусов одного и того же рода в кишечнике как здоровых, так и плохо функционирующих птиц указывает на существование разных патотипов вируса. Необходимы дальнейшие исследования, чтобы определить обоснованность этих утверждений.
С появлением молекулярных диагностических тестов и секвенирования наше понимание кишечных вирусов расширилось.Также существует вероятность того, что наши текущие методы не могут обнаружить некоторые еще неизвестные вирусы / патогены, которые могут иметь последствия для энтерита индейки. Доступность и использование высокопроизводительных методов пиросеквенирования нуклеиновых кислот нового поколения, таких как секвенирование Illumina, может выявить последовательности агентов, о которых ранее не сообщалось у индеек. Метагеномные исследования РНК-вирусов в кишечнике индеек, страдающих кишечным заболеванием, выявили присутствие последовательностей вирусов дцРНК ( Reoviridae и пикобирнавирусы) и вирусов оцРНК ( Caliciviridae , Leviviridae , Picornaviridae Astroviridae и Picornaviridae ). [18].Обнаружение неизвестных вирусов методом метагеномики проложит путь к разработке методов диагностики этих вирусов. Кроме того, необходимо определить, почему одни и те же типы вируса обнаруживаются в стадах индейки, пораженных энтеритом, а также в явно здоровых стадах. Полное понимание роли кишечных вирусов и других патогенов в кишечных болезнях индеек будет иметь большое значение при разработке профилактических и контрольных мер.
Ссылки
1. Акимкин В., Биндель Ф., Хоферер М., Стинг Р., Полли Б., Ханель А., Хафез Х.М.Одношаговая RT-qPCR с системой внутреннего контроля для обнаружения ротавирусов индейки в образцах фекалий. J Virol Methods. 2011. 177 (1): 112–117. [PubMed] [Google Scholar] 2. Асано К. М., Грегори Ф., Соуза С. П., Ротава Д., Оливейра Р. Н., Вильярреал Л. Я., Рихтценхайн Л. Дж., Брандао ЧП. Ротавирус крупного рогатого скота у индеек с энтеритом. Птичий Дис. 2011; 55 (4): 697–700. [PubMed] [Google Scholar] 3. Трепет О.О., Али А., Элайш М., Ибрагим М., Мурджа М., Пантин-Джеквуд М., Саиф Ю.М., Ли К.В. Влияние коронавирусной инфекции на репродуктивную способность кур индюков.Птичий Дис. 2013; 57: 650–656. [PubMed] [Google Scholar] 4. Баньяи К., Дандар Э., Дорси К.М., Мато Т., Паля В. Геномное созвездие нового штамма ортореовируса птиц, связанного с синдромом низкорослости у бройлеров. Гены вирусов. 2011; 42: 82–89. [PubMed] [Google Scholar]5. Барнс Х. Дж., Гай Дж. С.. Синдром энтерита-смертности птицы. В: Саиф Ю.М., Барнс Х.Дж., Глиссон-младший, Фэдли А.М., Макдугалд Л.Р., Суэйн Д.Е., редакторы. Болезни птицы. 11-е изд. Эймс: Издательство государственного университета Айовы; 2003. С. 1171–1180.
6. Barnes HJ, Guy JS, Vaillancourt JP. Комплекс энтерита птицы. Rev Sci Tech. 2000. 19: 565–588. [PubMed] [Google Scholar] 7. Бенавенте Дж., Мартинес-Костас Дж. Птичий реовирус: структура и биология. Virus Res. 2007. 123: 105–119. [PubMed] [Google Scholar] 8. Безерра Д.А., да Силва Р.Р., Кайано Дж. Х., Сильвестр Р. В., де Соуза Оливейра Д., Линьярес С., Габбай Ю. Б., Маскареньяс Дж. Д.. Обнаружение ротавируса группы D птиц с использованием полимеразной цепной реакции для гена VP6. J Virol Methods. 2012; 185: 189–192. [PubMed] [Google Scholar] 9.Bidin M, Bidin Z, Majnaric D, Tisljar M, Lojkic I. Циркуляция и филогенетическая взаимосвязь астровирусов куриного и индейского происхождения, обнаруженных у домашних уток ( Anas platyrhynchos domesticus ) Avian Pathol. 2012. 41 (6): 555–562. [PubMed] [Google Scholar] 10. Боделон Дж., Лабрада Л., Мартинез-Костас Дж., Бенавенте Дж. Сегмент генома реовируса птиц S1 представляет собой функционально трицистронный ген, который экспрессирует один структурный и два неструктурных белка в инфицированных клетках. Вирусология. 2001; 290: 181–191.[PubMed] [Google Scholar] 11. Бреслин Дж. Дж., Смит Л. Г., Барнс Г. Дж., Гай Дж. С.. Сравнение процедур выделения вируса, иммуногистохимии и полимеразной цепной реакции с обратной транскриптазой для обнаружения коронавируса индейки. Птичий Дис. 2000. 44: 624–631. [PubMed] [Google Scholar] 12. Бангер А. Н., Чакон Дж. Л., Джонс Р. К., Феррейра А. Дж.. Обнаружение и молекулярная характеристика гена 3 и 5 коронавируса индейки у индюшат с тяжелым энтеритом в Бразилии. Птичий Дис. 2009. 53: 356–362. [PubMed] [Google Scholar] 13.Канелли Э., Кордиоли П., Барбьери И., Кателла А., Пеннелли Д., Церути Р., Морено А., Лавацца А. Астровирусы как возбудители энтерита домашней птицы: генетическая характеристика и лонгитюдные исследования в полевых условиях. Птичий Дис. 2012; 56: 173–182. [PubMed] [Google Scholar] 14. Cardoso TC, Castanheira TL, Teixeira MC, Rosa AC, Hirata KY, Astolphi RD, Luvizotto MC. Валидация иммуногистохимического анализа для обнаружения коронавируса индейки: быстрый и простой инструмент для скрининга в условиях ограниченных ресурсов. Poult Sci.2008. 87 (7): 1347–1352. [Бесплатная статья PMC] [PubMed] [Google Scholar] 15. Чен Ю.Н., Ву СС, Брайан Т., Хупер Т., Шредер Д., Лин Т.Л. Специфическая полимеразная цепная реакция обратной транскрипции в режиме реального времени для обнаружения и количественного определения РНК коронавируса индейки в тканях и фекалиях индеек, инфицированных коронавирусом индейки. J Virol Methods. 2010. 163: 452–458. [Бесплатная статья PMC] [PubMed] [Google Scholar] 16. Калвер Ф.А., Бриттон П., Кавана Д. Обнаружение с помощью ОТ-ПЦР птичьих коронавирусов галлиформных птиц (курица, индейка, фазан) и у попугая.Методы Мол биол. 2008; 454: 35–42. [Бесплатная статья PMC] [PubMed] [Google Scholar] 17. Да Силва SEL, Бонетти AM, Petrocelli ATM, Ferrari HF, Luvizotto MCR, Cardoso TC. Обнаружение астровируса индейки у молодняка, пораженного комплексом энтерита домашней птицы, в Бразилии. J Vet Med Sci. 2008. 70 (6): 629–631. [PubMed] [Google Scholar]19. Дэй Дж. М., Гондер Э., Дженнингс С., Ривес Д., Тилли Б., Вуминг Б. Молекулярная характеристика кишечных коронавирусов индейки, циркулирующих в США в 2012 году. Документ, представленный на ежегодном собрании Американской ассоциации патологоанатома в Чикаго, штат Иллинойс.2013.
20. Day JM, Pantin-Jackwood MJ, Spackman E. Последовательный и филогенетический анализ сегмента S1 генома реовирусов индейского происхождения. Гены вирусов. 2007. 35 (2): 235–242. [PubMed] [Google Scholar] 21. Дэй JM, Spackman E, Пантин-Джеквуд MJ. Мультиплексный ОТ-ПЦР-тест для дифференциальной идентификации астровируса индейки типа 1, астровируса индейки типа 2, астровируса цыпленка, вируса птичьего нефрита и птичьего ротавируса. Птичий Дис. 2007. 51: 681–684. [PubMed] [Google Scholar] 22. Дэй JM, Spackman E, Пантин-Джеквуд MJ.Иммунная дисфункция, вызванная реовирусом индейского происхождения, у свободных от конкретных патогенов и промышленных индюшат. Птичий Дис. 2008. 52 (3): 387–391. [PubMed] [Google Scholar] 23. Де Баттисти С., Сальвиато А., Йонассен С.М., Тоффан А., Капуа I, Каттоли Г. Генетическая характеристика астровирусов, обнаруженных у цесарок ( Numida meleagris ), выявляет отдельный генотип и предполагает межвидовую передачу между индейкой и цесаркой. Arch Virol. 2012. 157 (7): 1329–1337. [PubMed] [Google Scholar] 24. Де Бенедиктис П., Шульц-Черри С., Бернем А., Каттоли Г.Астровирусные инфекции у людей и животных — молекулярная биология, генетическое разнообразие и межвидовая передача. Заразить Genet Evol. 2011. 11 (7): 1529–1544. [Бесплатная статья PMC] [PubMed] [Google Scholar] 25. Девитт CM, Рейнольдс DL. Характеристика ротавирусов группы D. Птичий Дис. 1993; 37: 749–755. [PubMed] [Google Scholar] 26. de Wit JJ, ten Dam GB, van de Laar JMAM, Biermann Y, Verstegen I, Edens F, Schrier CC. Обнаружение и характеристика нового астровируса у кур и индеек с кишечными расстройствами и нарушениями опорно-двигательного аппарата.Avian Pathol. 2011; 40: 453–461. [PubMed] [Google Scholar] 27. Доманска-Бличарц К., Серока А., Минта З. Годовое молекулярное исследование астровирусной инфекции у индеек в Польше. Arch Virol. 2011. 156 (6): 1065–1072. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Франка М., Креспо Р., Чин Р., Вулкок П., Шивапрасад Х.Л. Ретроспективное исследование миокардита, ассоциированного с реовирусом, у индеек. Птичий Дис. 2010. 54 (3): 1026–1031. [PubMed] [Google Scholar] 30. Gomaa MH, Yoo D, Ojkic D, Barta JR. Распространенность коронавируса индейки у североамериканских индеек определяется с помощью недавно разработанного иммуноферментного анализа на основе рекомбинантного антигена.Clin Vaccine Immunol. 2008. 15 (12): 1839–1844. [Бесплатная статья PMC] [PubMed] [Google Scholar] 31. Gomaa MH, Yoo D, Ojkic D, Barta JR. Использование рекомбинантного полипептида-шипа S1 для разработки ELISA-анализа антител, специфичных к TCoV. Vet Microbiol. 2009. 138 (3–4): 281–288. [Бесплатная статья PMC] [PubMed] [Google Scholar] 32. Gomaa MH, Yoo D, Ojkic D, Barta JR. Заражение патогенным изолятом коронавируса индейки отрицательно влияет на показатели роста и морфологию кишечника молодняка индейки в Канаде. Avian Pathol.2009. 38: 279–286. [PubMed] [Google Scholar] 33. Гомес Д.Е., Хирата К.Ю., Сахеки К., Роза А.С., Лувизотто М.С., Кардосо ТК. Патология и тканевое распределение коронавируса индейки у экспериментально инфицированных цыплят и индюшат. J Comp Pathol. 2010. 143 (1): 8–13. [Бесплатная статья PMC] [PubMed] [Google Scholar] 34. Гай JS. Коронавирусный энтерит в Турции. В: Саиф Ю.М., Барнс Х.Дж., Глиссон-младший, Фэдли А.М., Макдугалд Л.Р., Суэйн Д.Е., редакторы. Болезнь птицы. 11. Эймс: Издательство государственного университета Айовы; 2003. С. 300–307. [Google Scholar] 35.Гай JS. Коронавирусный энтерит в Турции. В: Саиф YM, Fadly AM, Glisson JR, McDougald LR, Nolan LK, Swayne DE, редакторы. Болезни птицы. 12. Эймс: специалист по издательскому делу Wiley-Blackwell; 2008. С. 330–338. [Google Scholar] 36. Guy JS, Miles AM, Smith L, Fuller FJ, Schultz-Cherry S. Антигенная и геномная характеристика энтеровирусоподобного вируса индейки (Северная Каролина, изолят 1988 г.): идентификация вируса как астровируса индейки 2. Avian Dis. 2004. 48 (1): 206–211. [PubMed] [Google Scholar] 37.Гай Дж. С., Смит Л. Г., Бреслин Дж. Дж., Пакпиньо С. Разработка конкурентного иммуноферментного анализа для обнаружения антител к коронавирусу индейки. Птичий Дис. 2002. 46 (2): 334–341. [PubMed] [Google Scholar] 38. Гай Дж. С., Смит Л. Г., Бреслин Дж. Дж., Вайланкур Дж. П., Барнс Х. Дж. Высокая смертность и подавление роста, экспериментально вызванные у молодых индеек двойным заражением энтеропатогенным вирусом Escherichia coli и коронавирусом индейки. Птичий Дис. 2000. 44: 105–113. [PubMed] [Google Scholar] 39.Heggen-Peay CL, Qureshi MA, Edens FW, Sherry B, Wakenell PS, O’Connell PH, Schat KA. Выделение реовируса от синдрома энтерита и смертности домашних птиц и его патогенность у индюшат. Птичий Дис. 2002. 46 (1): 32–47. [PubMed] [Google Scholar] 40. Имада Т., Ямагути С., Масе М, Цукамото К., Кубо М., Морока А. Вирус птичьего нефрита (ANV) как новый член семейства Astroviridae и конструирование инфекционной кДНК ANV. J Virol. 2000; 74: 8487–8493. [Бесплатная статья PMC] [PubMed] [Google Scholar] 41.Исмаил М.М., Тан А.Ю., Саиф Ю.М. Патогенность коронавируса индейки у индеек и кур. Птичий Дис. 2003. 47: 515–522. [PubMed] [Google Scholar] 42. Джеквуд М.У., Бойнтон Т.О., Хилт Д.А., Маккинли Е.Т., Киссинджер Дж.С., Патерсон А.Х., Робертсон Дж., Лемке С., МакКолл А.В., Уильямс С.М., Джеквуд Дж. В., Берд Л.А. Возникновение коронавируса группы 3 в результате рекомбинации. Вирусология. 2010; 398: 98–108. [Бесплатная статья PMC] [PubMed] [Google Scholar] 43. Джиндал Н., Чандер И., Патнаяк Д.П., Мор СК, Зиглер А.Ф., Гоял С.М. Мультиплексная RT-PCR для обнаружения астровирусов, ротавирусов и реовирусов у индеек.Птичий Дис. 2012; 56: 592–596. [PubMed] [Google Scholar] 44. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М. Выявление и молекулярная характеристика кишечных вирусов синдрома энтерита домашней птицы у индеек. Poult Sci. 2010. 89: 217–226. [Бесплатная статья PMC] [PubMed] [Google Scholar] 45. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М. Обнаружение и молекулярная характеристика кишечных вирусов у племенных индюков. Avian Pathol. 2010; 39: 53–61. [PubMed] [Google Scholar] 46. Джиндал Н., Патнаяк Д.П., Чандер И., Зиглер А.Ф., Гоял С.М.Сравнение последовательностей капсидных генов астровируса-2 индейки у индюшат, пораженных синдромом энтерита, и у практически здоровых индюшат. Arch Virol. 2011; 156: 969–977. [PubMed] [Google Scholar] 47. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Ретроспективное исследование синдрома энтерита домашней птицы в Миннесоте. Птичий Дис. 2009; 53: 268–275. [PubMed] [Google Scholar] 48. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Экспериментальное воспроизведение синдрома энтерита домашней птицы: клинические данные, реакция роста и микробиология.Poult Sci. 2009; 88: 949–958. [Бесплатная статья PMC] [PubMed] [Google Scholar] 49. Джиндал Н., Патнаяк Д.П., Зиглер А., Лаго А., Гоял С.М. Продолжительность угнетения роста и выделения патогенов при синдроме энтерита экспериментально воспроизведенных индюшат. Птичий Дис. 2009; 53: 517–522. [PubMed] [Google Scholar] 50. Johne R, Otto P, Roth B, Löhren U, Belnap D, Reetz J, Trojnar E. Анализ последовательности сегмента генома, кодирующего VP6, ротавирусов птичьей группы F и G. Вирусология. 2011; 412: 384–391. [PubMed] [Google Scholar] 51.Джотикумар Н, Канг Г., Хилл ВР. Широкореактивный анализ TaqMan для обнаружения ротавируса в режиме реального времени с помощью ОТ-ПЦР в клинических образцах и образцах окружающей среды. J Virol Methods. 2009. 155: 126–131. [PubMed] [Google Scholar] 52. Капчинский Д.Р., Селлерс Х.С., Симмонс В., Шульц-Черри С. Анализ последовательности гена S3 из реовируса индейки. Гены вирусов. 2002. 25 (1): 95–100. [Бесплатная статья PMC] [PubMed] [Google Scholar] 53. Кэ GM, Cheng HL, Ke LY, Ji WT, Chulu JL, Liao MH, Chang TJ, Liu HJ. Разработка количественного светового циклера RT-PCR в реальном времени для обнаружения птичьего реовируса.J Virol Methods. 2006; 133 (1): 6–13. [PubMed] [Google Scholar] 54. King AMQ, Lefkowiz E, Adams M, Carstens E. Таксономия вирусов: девятый доклад международного комитета по таксономии вирусов. Уолтем: Academic Press; 2012. [Google Scholar] 55. Koci MD, Schultz-Cherry S. Птичьи астровирусы. Avian Pathol. 2002; 31: 213–227. [PubMed] [Google Scholar] 56. Koci MD, Seal BS, Schultz-Cherry S. Разработка диагностического теста RT-PCR для птичьего астровируса. J Virol Methods. 2000. 90: 79–83. [Бесплатная статья PMC] [PubMed] [Google Scholar] 58.Кул Д.А., Холмс И.Х. Нуклеотидная и расчетная аминокислотные последовательности Ty-1 Vp7 ротавируса птиц значительно отличаются от таковых ротавируса Ch-2. Arch Virol. 1993. 129 (1–4): 227–234. [PubMed] [Google Scholar] 59. Lin TL, Loa CC, Tsai SC, Wu CC, Bryan TA, Thacker HL, Hooper T., Schrader D. Характеристика коронавируса индейки у индюшат с острым энтеритом. Vet Microbiol. 2002. 84 (1–2): 179–186. [Бесплатная статья PMC] [PubMed] [Google Scholar] 60. Линь Т.Л., Лоа СС, Ву СС. Полные последовательности 3′-концевой кодирующей области для генов структурных белков коронавируса индейки.Virus Res. 2004. 106 (1): 61–70. [Бесплатная статья PMC] [PubMed] [Google Scholar] 61. Loa CC, Wu CC, Lin TL. Сравнение 3′-концевых кодирующих областей изолятов коронавируса индейки из Индианы, Северной Каролины и Миннесоты со штаммами коронавируса инфекционного бронхита кур. Интервирология. 2006. 49 (4): 230–238. [Бесплатная статья PMC] [PubMed] [Google Scholar] 62. Lojkic I, Bidin M, Bidin Z, Mikec M. Вирусные агенты, связанные с энтеритом домашней птицы в хорватских промышленных стадах индейки. Acta Vet BRNO. 2010; 79: 91–98.[Google Scholar] 63. Маттейнссенс Дж., Мартелла В., Ван Ранст М. Оценка приоритетной статьи: геномная эволюция, межвидовой барьер хозяина, реассортировка и классификация ротавирусов. Будущий Virol. 2010. 5: 385–390. [Google Scholar] 64. Matthijnssens J, Otto PH, Ciarlet M, Desselberger U, Van Ranst M, Johne R. Значения отсечения на основе последовательности VP6 в качестве критерия для демаркации видов ротавирусов. Arch Virol. 2012. 157 (6): 1177–1182. [PubMed] [Google Scholar] 65. Maurel S, Toquin D, Briand FX, Queguiner M, Allee C, Bertin J, Ravillion L, Retaux C, Turblin V, Morvan H, Eterradossi N.Первые полноразмерные последовательности гена S европейских изолятов выявили дальнейшее разнообразие коронавирусов индейки. Avian Pathol. 2011; 40: 179–189. [PubMed] [Google Scholar] 66. Макналти М.С., Аллан Г.М., Маккракен Р.М. Экспериментальное заражение цыплят ротавирусами: клинические и вирусологические данные. Avian Pathol. 1983; 12: 45–54. [PubMed] [Google Scholar] 67. Мор С.К., Абин М., Коста Дж., Дуррани А., Джиндал Н., Гоял С.М., Патнаяк Д.П. Роль астровируса индейки 2 типа в синдроме энтерита домашних птиц. Poult Sci.2011; 90: 2747–2752. [Бесплатная статья PMC] [PubMed] [Google Scholar] 68. Мор С.К., Шарафельдин Т.А., Абин М., Кромм М., Портер Р.Э., Гоял С.М., Патнаяк Д.П. Возникновение кишечных вирусов при синдроме легкой индейки. Avian Pathol. 2013. 42 (5): 497–501. [PubMed] [Google Scholar] 69. Мор С.К., Шарафельдин Т.А., Портер Р.Э., Зиглер А., Патнаяк Д.П., Гоял С.М. Выделение и характеристика реовируса артрита индейки. Птичий Дис. 2013. 57 (1): 97–103. [PubMed] [Google Scholar] 70. Моура-Альварес Дж., Чакон СП, Сканавини Л.С., Нуньес ЛФН, Астольфи-Феррейра К.С., Джонс Р.К., Пиантино Феррейра А.Дж.Кишечные вирусы в стадах бразильских индейок: частота единичных и множественных вирусных инфекций в зависимости от возраста и клинических признаков кишечного заболевания. Poult Sci. 2013; 92: 945–955. [Бесплатная статья PMC] [PubMed] [Google Scholar] 71. Otto PH, Ahmed MU, Hotzel H, Machnowska P, Reetz J, Roth B, Trojnar E, Johne R. Обнаружение птичьих ротавирусов групп A, D, F и G у больных кур и индеек из Европы и Бангладеш. Vet Microbiol. 2012; 156 (1-2): 8-15. [Бесплатная статья PMC] [PubMed] [Google Scholar] 72.Отто П., Либлер-Тенорио Е.М., Эльшнер М., Ритц Дж., Лорен У., Диллер Р. Обнаружение ротавирусов и поражений кишечника у цыплят-бройлеров в стадах с синдромом короткого роста и задержки роста (RSS) Avian Dis. 2006; 50: 411–418. [PubMed] [Google Scholar] 73. Пакпиньо С., Лей Д.Х., Барнс Х.Дж., Вайланкур Дж. П., Гай Дж. С.. Распространенность энтеропатогенной Escherichia coli в естественных случаях синдрома энтерита-смертности домашних птиц. Птичий Дис. 2002. 46 (2): 360–369. [PubMed] [Google Scholar] 74. Palade EA, Demeter Z, Hornyak A, Nemes C, Kisary J, Rusvai M.Высокая распространенность парвовируса индейки в стадах индейки из Венгрии, страдающих синдромами кишечных заболеваний. Птичий Дис. 2011; 55 (3): 468–475. [PubMed] [Google Scholar] 75. Пантин-Джеквуд MJ, Day JM, Jackwood MW, Spackman E. Кишечные вирусы, обнаруженные молекулярными методами в коммерческих стадах кур и индейок в Соединенных Штатах в период с 2005 по 2006 гг. Птичий Дис. 2008. 52: 235–244. [PubMed] [Google Scholar] 76. Пантин-Джеквуд MJ, Spackman E, Day JM. Патогенез астровирусов индейки 2 типа с вариантными генами капсида у 2-дневных цыплят, свободных от специфических патогенов.Avian Pathol. 2008; 37: 193–201. [PubMed] [Google Scholar] 77. Пантин-Джеквуд MJ, Spackman E, Day JM, Rives D. Периодический мониторинг промышленных индеек на предмет энтеросолюбии указывает на постоянное присутствие астровируса и ротавируса на фермах. Птичий Дис. 2007. 51: 674–680. [PubMed] [Google Scholar] 78. Пантин-Джеквуд MJ, Spackman E, Woolcock PR. Молекулярная характеристика и типирование астровирусов кур и индейки, циркулирующих в США: значение для диагностики. Птичий Дис. 2006; 50: 397–404.[PubMed] [Google Scholar] 79. Пантин-Джеквуд MJ, Strother KO, Mundt E, Zsak L, Day JM, Spackman E. Молекулярная характеристика птичьих астровирусов. Arch Virol. 2011; 156: 235–244. [PubMed] [Google Scholar] 80. Куреши М.А., Саиф Ю.М., Хегген-Пи К.Л., Эденс Ф.В., Хавенштейн ГБ. Индукция функциональных дефектов в макрофагах астровирусом индейки, ассоциированным с энтеритом и синдромом смертности домашних птиц. Птичий Дис. 2001. 45 (4): 853–861. [PubMed] [Google Scholar] 81. Рамиг РФ, Ciarlet M, Mertens PPC, Dermody TS. Ротавирус.В: Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA, редакторы. Таксономия вирусов: восьмой отчет международного комитета по таксономии вирусов. Амстердам: Academic Press; 2005. С. 484–496. [Google Scholar] 82. Рейнольдс Д.Л., Саиф Ю.М. Астровирус: причина кишечного заболевания у индюшат. Птичий Дис. 1986; 30: 728–735. [PubMed] [Google Scholar]83. Роббинс К.М. Эпидемиология и диагностика коронавируса в Турции. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс.2013.
84. Родригес-Диас Дж., Рубилар-Абреу Э., Шпицнер М., Хедлунд К.О., Липранди Ф., Свенссон Л. Дизайн мультиплексной вложенной ПЦР для генотипирования NSP4 ротавируса группы А. J Virol Methods. 2008. 149: 240–245. [PubMed] [Google Scholar] 85. Ruff MD, Rosenberger JK. Сопутствующие инфекции реовирусами и кокцидиями у бройлеров. Птичий Дис. 1985. 29 (2): 465–478. [PubMed] [Google Scholar] 86. Шульц-Черри С., Капчинский Д.Р., Симмонс В.М., Кочи М.Д., Браун С., Барнс Х.Дж. Идентификация агента (ов), связанного с синдромом смертности от энтерита домашней птицы: важность тимуса.Птичий Дис. 2000. 44: 256–265. [PubMed] [Google Scholar] 87. Шуман Т., Хотцель Х., Отто П., Джон Р. Доказательства межвидовой передачи и реассортации среди ротавирусов птичьей группы А. Вирусология. 2009. 386 (2): 334–343. [PubMed] [Google Scholar] 88. Продавцы HS, Linnemann EG, Pereira L, Kapczynski DR. Филогенетический анализ гена белка сигма 2 реовирусов индейки. Птичий Дис. 2004. 48 (3): 651–657. [PubMed] [Google Scholar] 89. Продавцы HS, Koci MD, Linnemann E, Kelley LA, Schultz-Cherry S. Разработка диагностического теста мультиплексной полимеразной цепной реакции с обратной транскрипцией, специфичного для астровируса и коронавируса индейки.Птичий Дис. 2004. 48: 531–539. [PubMed] [Google Scholar] 90. Shivaprasad HL, Franca M, Woolcock PR, Nordhausen R, Day JM, Pantin-Jackwood M. Миокардит, связанный с реовирусом у индюшат. Птичий Дис. 2009. 53: 523–532. [PubMed] [Google Scholar] 91. Spackman E, Day JM, Pantin-Jackwood MJ. Сопутствующие инфекции астровируса, реовируса и ротавируса вызывают снижение прибавки в весе у широкогрудых белых цыплят. Птичий Дис. 2010; 54: 16–21. [PubMed] [Google Scholar] 92. Спакман Э., Капчинский Д., Селлерс Х. Мультиплексная полимеразная цепная реакция обратной транскрипции в реальном времени для обнаружения трех вирусов, связанных с комплексом энтерита домашней птицы: астровируса индейки, коронавируса индейки и реовируса индейки.Птичий Дис. 2005; 49: 86–91. [PubMed] [Google Scholar] 93. Спакман Э., Пантин-Джеквуд М., Дэй Дж. М., Селлерс Х. Патогенез реовирусов индейского происхождения у индеек и кур. Avian Pathol. 2005; 34: 291–296. [PubMed] [Google Scholar] 94. Штамм E, Kelley LA, Schultz-Cherry S, Muse SV, Koci MD. Геномный анализ близкородственных астровирусов. J Virol. 2008. 82: 5099–5103. [Бесплатная статья PMC] [PubMed] [Google Scholar] 95. Сугияма М.К., Гото К., Уэмукай Х., Мори Й., Ито Н., Минамото Н. Присоединение и инфицирование к клеткам MA104 ротавирусов птиц требует присутствия сиаловой кислоты на поверхности клетки.J Vet Med Sci. 2004. 66: 461–463. [PubMed] [Google Scholar] 96. Тан Й., Исмаил М.М., Саиф Ю.М. Разработка иммуноферментного анализа с захватом антигена и ОТ-ПЦР для обнаружения астровирусов индейки. Птичий Дис. 2005. 49 (2): 182–188. [PubMed] [Google Scholar] 97. Тан Й., Мурджа А.М., Саиф Ю.М. Молекулярная характеристика гена капсида двух серотипов астровирусов индейки. Птичий Дис. 2005. 49 (4): 514–519. [PubMed] [Google Scholar] 98. MC Teixeira, MC Luvizotto, Ferrari HF, Mendes AR, da Silva SE, Cardoso TC.Обнаружение коронавируса индейки у коммерческих индюшат в Бразилии. Avian Pathol. 2007. 36 (1): 29–33. [PubMed] [Google Scholar] 99. Тейл К.В., Саиф Ю.М. Возрастные инфекции ротавируса, ротавирусоподобного вируса и атипичного ротавируса в стадах индейки. J Clin Microbiol. 1987. 25 (2): 333–337. [Бесплатная статья PMC] [PubMed] [Google Scholar]100. Тилли Б.Дж., Гондер Э., Мейсон С., Уикс Дж. Вспышка коронавируса в Турции в Северной Каролине. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс.2013.
101. Тоффан А., Катания С., Сальвиато А., Де Баттисти С., Васкеллари М., Тосон М., Капуа I, Каттоли Г. Экспериментальное заражение домашних птиц и цесарок генетически отличными птичьими астровирусами. Avian Pathol. 2012; 41: 429–435. [PubMed] [Google Scholar] 102. Trojnar E, Otto P, Roth B, Reetz J, Johne R. Сегменты генома ротавируса группы D имеют консервативные концы группы A, но кодируют специфичные для группы белки. J Virol. 2010; 84: 10254–10265. [Бесплатная статья PMC] [PubMed] [Google Scholar] 103.Trojnar E, Sachsenroder J, Twardziok S, Reetz J, Otto PH, Johne R. Идентификация ротавируса птичьей группы А, содержащего новый ген VP4, близкий родству к генам ротавирусов млекопитающих. J Gen Virol. 2013; 94: 136–142. [PubMed] [Google Scholar] 104. Велаюдхан Б.Т., Шин Х.Д., Лопес В.К., Хупер Т., Халворсон Д.А., Нагараджа К.В. Анализ обратной транскриптазы-полимеразной цепной реакции для диагностики коронавирусной инфекции индейки. J Vet Diagn Invest. 2003. 15 (6): 592–596. [PubMed] [Google Scholar] 105.Woolcock PR, Shivaprasad HL. Электронно-микроскопическая идентификация вирусов, связанных с энтеритом домашней птицы, у индеек, выращенных в Калифорнии в 1993–2003 гг. Птичий Дис. 2008. 52: 209–213. [PubMed] [Google Scholar]106. Вуминг Б. Вспышка коронавируса на северо-западе Арканзаса. Документ представлен на ежегодном собрании Американской ассоциации патологов птиц, проходившем в Чикаго, штат Иллинойс. 2013.
107. Ю М., Исмаил М. М., Куреши М. А., Дерт Р. Н., Барнс Х. Дж., Саиф Ю. М.. Вирусные агенты, связанные с энтеритом и синдромом смертности домашних птиц: роль небольшого круглого вируса и коронавируса индейки.Птичий Дис. 2000. 44 (2): 297–304. [PubMed] [Google Scholar] 108. Ю М, Тан Й, Го М, Чжан Кью, Саиф Ю. Характеристика небольшого круглого вируса, связанного с синдромом энтерита и смертности цыплят. Птичий Дис. 2000. 44 (3): 600–610. [PubMed] [Google Scholar] 109. Zsak L, Strother KO, Kisary J. Анализ частичной последовательности генома парвовирусов, связанных с кишечным заболеванием у домашней птицы. Avian Pathol. 2008. 37 (4): 435–441. [PubMed] [Google Scholar]вирусов для Турции | Дикая природа | Департамент рыбы и дичи Нью-Гэмпшира
Выберите изображения для большего размера
Вирус птичьей оспы и лимфопролиферативного заболевания (LPDV)
В популяции дикой индейки присутствуют два вируса, которые могут вызывать видимые бородавчатые наросты или поражения на голове, шее или других неперых частях птицы:
Вирус птичьей оспы был обнаружен в США.S. на протяжении десятилетий, а вирус лимфопролиферативного заболевания впервые был зарегистрирован в США в 2009 году.
New Hampshire Fish and Game отслеживает распространенность у индюков с симптомами с 2011 года. Хотя из всех частей штата были получены многочисленные сообщения о птицах, проявляющих физические симптомы, эти вирусы, по-видимому, не оказывают негативного воздействия на популяцию индейки. в гос.
Первые две пораженные индейки были собраны в North Haverhill, N.H. в октябре 2011 года. В 2011 и 2012 годах наблюдались больные индейки, собирали образцы и отправляли их в государственные ветеринарные лаборатории в Нью-Гэмпшире. В некоторых из отправленных на анализ образцов индейки был обнаружен вирус как LPDV, так и вирус птичьей оспы.
В 2013 году охотники из Нью-Гэмпшира пожертвовали ножки выловленных индеек, которые были отправлены в Национальную лабораторию болезней дикой природы в Джорджии. Было обнаружено, что примерно 71% из них были носителями LPDV, но только у небольшого числа индеек действительно наблюдались физические симптомы.
Нет никаких известных последствий для здоровья человека, связанных ни с LPDV, ни с птичьей оспой. Вирусы не вредны и не передаются человеку.
СлужбаNew Hampshire Fish and Game продолжает отслеживать распространенность индеек с симптомами заболевания с помощью двух ежегодных онлайн-опросов. Просим всех, кто видит индюков, демонстрирующих физические признаки этих вирусов, сообщить о своих наблюдениях здесь:
Вирусный гепатит индейки
Чо Б.Р., 1976.Аденовирус индейки, патогенный как для цыплят, так и для индюшат. Болезни птиц, 20: 714-723.
Guy JS, 1997. Вирусный гепатит Турции. В: Calnek B, Barnes HJ, Beard CW, McDougald LR, Saif YM, ред. Болезни домашней птицы, издание 10. Эймс, Айова, США: Издательство государственного университета Айовы, 773-777.
Король; AMQ; Коричневый F; Кристиан П; Hovi T; Hyypia T; Ноулз, штат Нью-Джерси; Лимонный СМ; Незначительный PD; Palmenberg AC; Скерн Т; Stanway G, 2000. Picornaviridae. В: Regenmortel M, Fauquet C, Bishop D, Carstens E, Estes M, Lemon S, Maniloff J, Mayo M, McGeoch D, Pringle C, Wickner R, eds.Таксономия вирусов: седьмой отчет Международного комитета по таксономии вирусов. Academic Press, Сан-Диего, США, 657-678.
Кляйн PN; Castro AE; Meteyer CU; Рейнольдс Б; Swartzman-Andert JA; Cooper G; Подбородок RP; Shivaprasad HL, 1991. Экспериментальная передача вирусного гепатита индейки суточным домашним птицам и идентификация ассоциированных вирусных частиц, напоминающих пикорнавирусы. Болезни птиц, 35 (1): 115-125; 45 исх.
Mandelli GA; Ринальди А; Cervio G, 1966. Макроскопические и ультрамикроскопические поражения при гепатопанкреатите у индеек.Atti Soc Ital Sci Vet, 20: 541-545.
McDonald JWC; Randall CJ; Dagless MD, 1982. Пикорнавирусоподобный вирус, вызывающий гепатит и панкреатит у индеек. Ветеринарный журнал, 111: 323.
Mongeau JD; Truscott RB; Фергюсон А.Е .; Коннелл М.С., 1959. Вирусный гепатит у индеек. Болезни птиц, 3: 388-396.
Snoeyenbos GH, 1991. Вирусный гепатит в Турции. Болезни домашней птицы., Изд. 9: 699-701; 13 исх.
Snoeyenbos GH; Basch HI, 1960. Дальнейшие исследования вирусного гепатита у индеек.Болезни птиц, 4: 477-485.
Snoeyenbos GH; Basch HI; Севоян М., 1959. Инфекционный возбудитель гепатита у индеек. Болезни птиц, 3: 377-388.
Tzianabos T; Snoeyenbos GH, 1965a. Некоторые физико-химические свойства вируса гепатита индейки. Болезни птиц, 9: 152-156.
Tzianabos T; Snoeyenbos GH, 1965b. Клинические, иммунологические и серологические наблюдения за вирусным гепатитом индейки. Болезнь птиц, 9: 578-595.
Van der Heide L; Брустолон М; Лоусон М.Г., 1980.Патогенность для цыплят реовируса, выделенного от индеек. Болезнь птиц, 24: 989-997.
Заработная плата DP; Ficken MD, 1989. Криптоспоридиоз и вирусный гепатит индейки у индеек. Болезни птиц, 33 (1): 191-194; 22 исх.
Wilcock BP; Thacker HL, 1976. Очаговый некроз печени у индеек с геморрагическим энтеритом. Болезни птиц 20: 205-208.
Ссылки на распространение
Кляйн П. Н., Кастро А. Е., Метейер К. У., Рейнольдс Б., Шварцман-Андерт Д. А., Купер Г., Чин Р. П., Шивапрасад Н. Л., 1991.Экспериментальная передача вирусного гепатита индейки суточным домашним птицам и идентификация ассоциированных вирусных частиц, напоминающих пикорнавирусы. Болезни птиц. 35 (1), 115-125. DOI: 10.2307 / 15
Mandelli G, Rinaldi A, Cervio G, 1966. Крупные и ультрамикроскопические поражения при гепатопанкреатите у индеек. Atti Soc. курсив. Sci. ветеринар. 541-545.
McDonald JWC, Randall CJ, Dagless MD, 1982. Пикорнавирусоподобный вирус, вызывающий гепатит и панкреатит у индеек. В: Ветеринарный журнал, 111 323.
Монжо Дж. Д., Траскотт Р. Б., Фергюсон А. Е., Коннелл М. С., 1959. Вирусный гепатит у индеек. Болезни птиц. 388-396. DOI: 10.2307 / 1587578
Snoeyenbos G H, Basch H. I., Sevoian M, 1959. Инфекционный агент, вызывающий гепатит у индеек. Болезни птиц. 377-388. DOI: 10.2307 / 1587577
Заработная плата D P, Ficken M. D., 1989. Криптоспоридиоз и вирусный гепатит индейки у индеек. Болезни птиц. 33 (1), 191–194. DOI: 10.2307 / 15
(PDF) Текущий сценарий вирусных заболеваний в птицеводстве Турции
ALEXANDER, D.Дж. И СЕНН, Д.А. (2008) Болезнь Ньюкасла и другие птичьи парамиксовирусы. В:
Dufour-Zavala L, ed. Лабораторное руководство по изоляции, идентификации и характеристике птичьих патогенов
. 4-е изд. Афины, Джорджия: Американская ассоциация патологов птиц, 135–141.
ALEXANDER, D. (2003) Болезнь Ньюкасла, другие птичьи парамиксовирусы и пневмовирусные инфекции, in:
SAIF, Y.M., BARNES, H.J., GLISSON, J.R., FADLY, A.M., MCDOUGALD, L.R. И СВАЙН, Д.(Ред.)
Болезни домашней птицы, стр. 63-99 (Эймс, издательство государственного университета Айовы).
ALFONSO-MORALES, A., MARTÍNEZ-PÉREZ, O., DOLZ, R., VALLE, R., PERERA, CL,
BERTRAN, K., FRÍAS, MT, MAJ, N., GANGES, L. и PÉREZ, LJ (2013) Spatiotemporal
Филогенетический анализ и молекулярная характеристика вирусов инфекционных заболеваний бурсальной полости на основе гипервариабельной области
VP2. PloS One 8: e65999.
ALKIE, T.N. и RAUTENSCHLEIN, S. (2016) Вирус инфекционной бурсальной болезни домашней птицы: текущее состояние и перспективы
на будущее.Ветеринарная медицина 7: 9-18.
АНОНИМНЫЙ (2014) Министерство продовольствия, сельского хозяйства и животноводства Турции, 2016. Годовой отчет,
Анкара.
ASHRAF, M.U., MAHMOOD, M.S., RAFIQUE, A., ABBAS, R.Z., IQBAL, Z., YOUNAS, M., SADIQ, S.
A., USMAN, M., ASGHAR, M.O. и ISHAQ, M.U. (2017) Факторы, ответственные за постоянную персистентность и эволюцию низкопатогенного вируса гриппа птиц (H9N2). World’s Poultry Science
Journal 73: 791-802.
BAYON-AUBOYER, MH, ARNAULD, C., TOQUIN, D. и ETERRADOSSI, N. (2000) Нуклеотидные
последовательности генов белков F, L и G двух птичьих пневмовирусов, отличных от A / не B ( APV) обнаруживают новую подгруппу
APV. Журнал общей вирусологии 81: 2723-2733.
BAYRAKTAR, E., UMAR, S., YILMAZ, AN, FRANZO, G., TUCCIARONE, CM, CECCHINATO,
M., CAKAN, B., IQBAL, M. и YILMAZ, H. (2018) Первый Молекулярная характеристика метапневмовируса птиц
(aMPV) в турецких бройлерных стадах.Болезнь птиц, 62: 425-430.
BIGGS, P.M. и НАИР, В. (2012) Долгосрочная перспектива: 40 лет исследований болезни Марека и птичьей патологии.
Патология птиц 41: 3-9.
BOODHOO, N., GURUNG, A., SHARIF, S. и BEHBOUDI, S. (2016) Болезнь Марека у кур: обзор
с акцентом на иммунологию. Ветеринарные исследования 47: 119.
БОЙНУКАРА, Б., ГУЛХАН, Т., КОВЕН, Ф., КИЗИРОГЛУ, И. и DURMU, A. (2013) Определение
вируса болезни Ньюкасла среди популяций диких птиц в бассейне озера Ван, Турция.Турецкий журнал
Ветеринария и зоотехника 37: 86-93.
КАВАНАХ, Д. (2007) Коронавирус птичьего инфекционного бронхита. Ветеринарные исследования 38: 281-297.
CECCHINATO, M., CATELLI, E., LUPINI, C., RICCHIZZI, E., CLUBBE, J., BATTILANI, M. и
NAYLOR, CJ (2010) Участие белка прикрепления птичьего метапневмовируса (AMPV) в вероятных вирус
эволюция одновременно с массовым внедрением живой вакцины. Ветеринарная микробиология 146: 24-34.
CECCHINATO, M., CATELLI, E., LUPINI, C., RICCHIZZI, E., PROSPERI, S. и NAYLOR, CJ
(2014) Возвращение вирулентности вакцины против птичьего метапневмовируса подтипа B: пришло время для регуляторы к
требуют наличия предшественников вакцины? Vaccine 32: 4660-4664.
ЧАКОН, J.L., FERREIRA, A.J.P. (2009) Дифференциация полевых изолятов и вакцинных штаммов
вируса инфекционного ларинготрахеита путем секвенирования ДНК. Vaccine 27: 6731-6738.
ЦИБЛАК, М.A., HASOKSUZ, M., ESCURET, V., VALETTE, M., GUL, F., YILMAZ, H., TURAN, N.,
BOZKAYA, E. and BADUR, S. (2009). Эпиднадзор и устойчивость к осельтамивиру вируса гриппа человека
в Турции в сезоне 2007-2008 гг. Журнал медицинской вирусологии 81: 1645-1651.
COOK, J.K.A., HUGGİNS, M.B., ORBELL, S.J. и SENNE, D.A, (1999) Предварительная антигенная характеристика
птичьего пневмовируса, выделенного от промышленных индеек в Колорадо, США. Птичий
Патология 28: 607-610.
COURTNEY, S.C., SUSTA, L., GOMEZ, D., HINES, N.L., PEDERSEN, J.C., BROWN, C.C., MILLER,
,P.J. and AFONSO, C.L. (2012) Сильно расходящиеся вирулентные изоляты вируса болезни Ньюкасла из Доминиканской Республики
являются членами нового генотипа, который, возможно, оставался незамеченным на протяжении более двух десятилетий.
Журнал клинической микробиологии 51: 508-517.
COVEN, F., RUTH MANVELL, ORHAN, G., COVEN, N. и GENC, A. (2004) YumurtacıVe Broyler
Surulerde Gorulen Newcastle Hastalığı Olgularından Avian Paramyxoviruslarınİzolasyonu Ve
VI. Ulusal Veteriner Mikrobiyoloji Kongresi. Kongre Ozet Kitabı, 222DAKMAN, A., GULEC, M., GUNAYDIN, E. and COŞAR, M. (2008) Патотипирование вируса болезни Ньюкасла
штаммов вируса, выделенных от домашних и диких птиц. Etlik Veterinary Mikrobiology Dergesi 19: 19-26.
DAKMAN, A., MUŞTAK, K., KARDOAN, O., DİVANOLU, Y., BİLGEN, N., AKAN, M. (2014)
Turkiye’de 2013 yılında izole edilen Newcastle Viruslarının Biyolojik,
Tiplendirilmesi.XI. Ветеринар Ulusal Hekimleri Mikrobiyoloji Kongresi, 21-24 Eylul Antalya.
FAOSTAT (2016) Статистическая база данных ФАО, по состоянию на июль 2016 года.
FENG, M. и ZHANG, X. (2016) Иммунитет к вирусу птичьего лейкоза: где мы сейчас и что делать
? Границы иммунологии 7: 624 DOI: 10.3389 / fi mmu.2016.00624.
World’s Poultry Science Journal, Vol. 75, декабрь 2019 г. 17
Вирусные болезни турецкой птицы: E. Bayraktar et al.
https: // www.cambridge.org/core/terms. https://doi.org/10.1017/S00439339106
Загружено с https://www.cambridge.org/core. Университет Флориды, 20 октября 2019 г.
