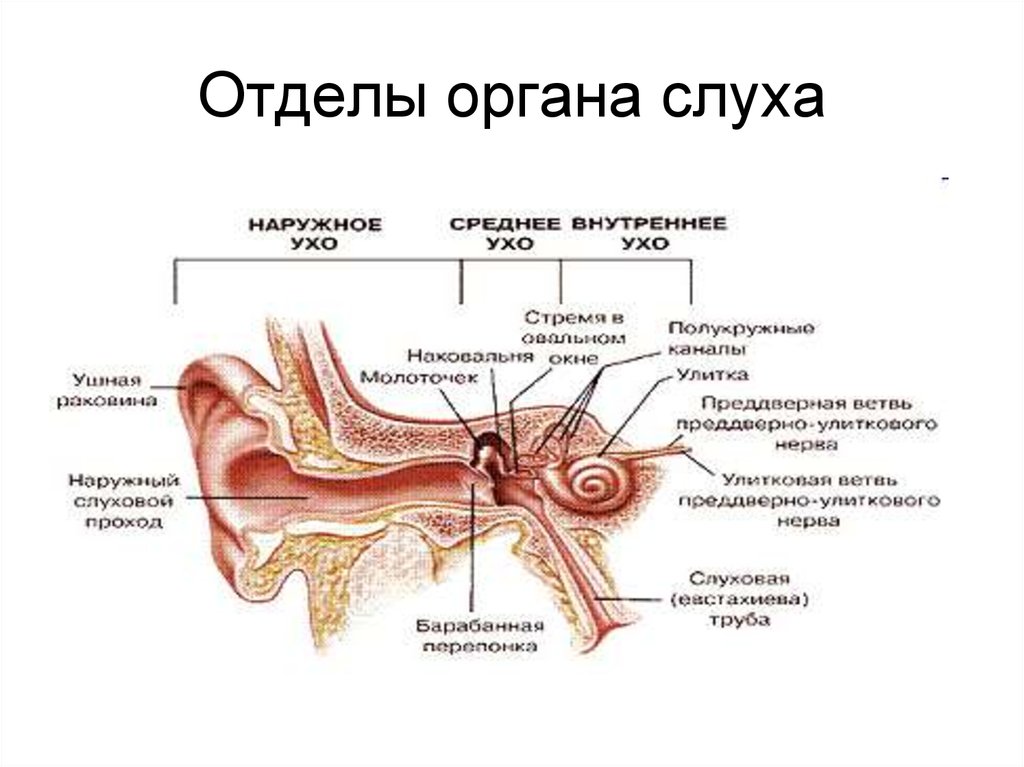
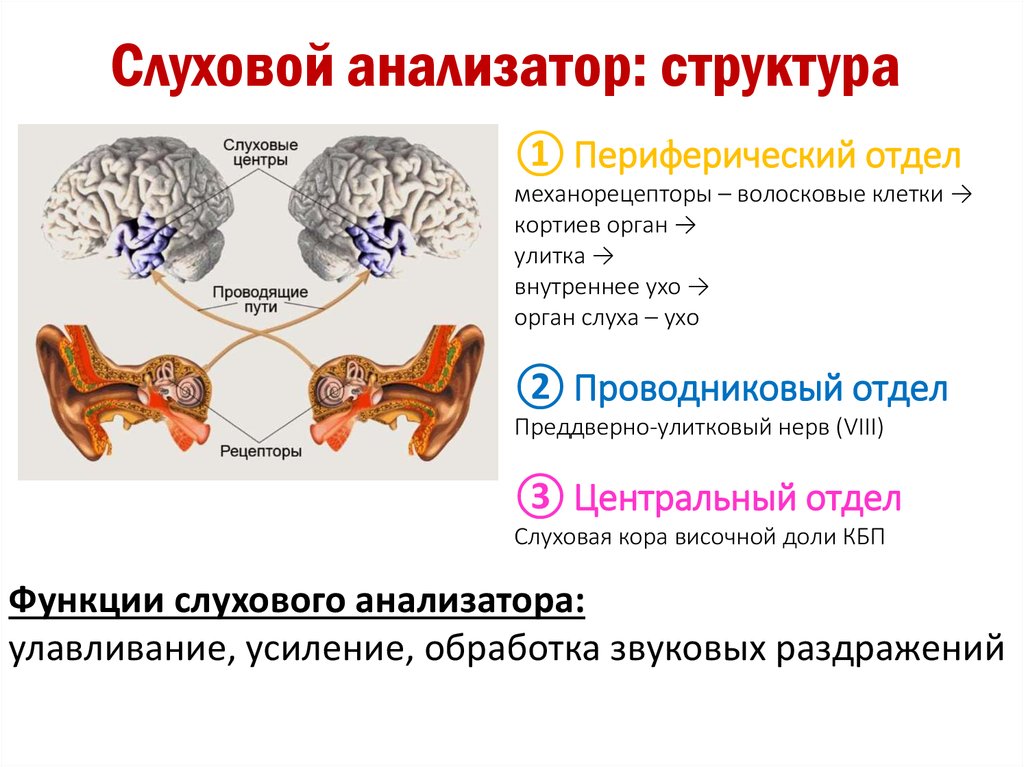
Как устроен орган слуха (слуховой анализатор): строение, функции
Воздушное и костное звукопроведение
Звуковая энергия поступает к структурам внутреннего уха путем воздушного звукопроведения и костного звукопроведения.
Воздушное звукопроведение – обычный путь поступления звуковых колебаний в ухо – через ушную раковину и наружный слуховой проход звук приходит к барабанной перепонке. Далее, колебания барабанной перепонки через цепь слуховых косточек передаются жидкостям слуховой улитки – пери- и эндолимфе, приводят в колебательное состояние основную мембрану и структуры кортиева органа.
Костное звукопроведение – это проведение звуковой вибрации от поверхности головы прямо в улитку внутреннего уха, минуя среднее ухо. При поступлении звуков в ухо путем костного звукопроведения звуковые колебания распространяются по костям и тканям головы. Под воздействием костнопроведенных звуков происходит вибрация стенок улитки внутреннего уха, которая передается наполняющим ее жидкостям. Это, в свою очередь, вызывает колебательные движения базилярной мембраны и кортиева органа. Далее все происходит так же, как при воздушном звукопроведении.
Это, в свою очередь, вызывает колебательные движения базилярной мембраны и кортиева органа. Далее все происходит так же, как при воздушном звукопроведении.
Собственный голос мы слышим именно посредством костного звукопроведения: звуки голоса проходят к улитке внутреннего уха через ткани головы. Именно поэтому мы слышим свой голос иначе, чем в записи. Это вызвано тем, что кости черепа проводят низкие частоты лучше, чем высокие. Поэтому во время звукопроизношения люди воспринимают собственный голос более низким и глубоким, чем его воспринимают окружающие.
Поскольку костное звукопроведение практически исключает среднее ухо из процесса передачи звука, то исследование слухового восприятия воздушно- и костнопроведенных звуков при проведении аудиометрии является очень важным при диагностике слуха.
Кроме того, в случаях невозможности слухопротезирования по воздушному звукопроведению, в частности, при определенных заболеваниях и после некоторых операций на среднем ухе, врач рассматривает возможность слухопротезирования по костному звукопроведению.
Промежуточный (проводниковый) отдел органа слуха
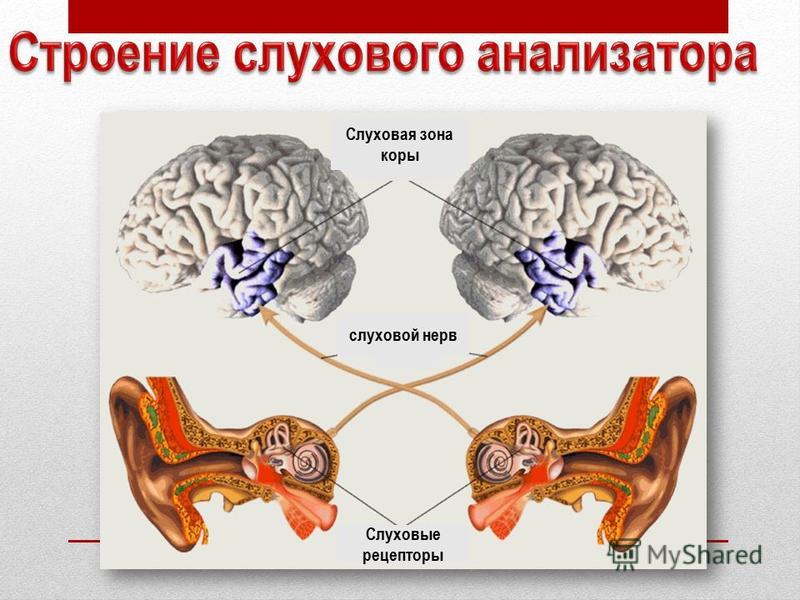
Промежуточный (проводниковый) отдел органа слуха начинается со слухового нерва и заканчивается в коре головного мозга. Тела нейронов слухового нерва расположены спирально по оси улитки и образуют так называемый спиральный ганглий. А их длинные отростки – аксоны – образуют слуховой нерв, передающий нервные импульсы «наверх» в мозг. Правый и левый слуховые нервы получили название восьмой (VIII) пары черепно-мозговых нервов.
Аксоны слухового нерва, как и других нейронов, покрыты слоем особой ткани – миелиновой оболочкой, в которой есть «перехваты» – оголенные участки аксона. Эта оболочка и ее «перехваты» играют ключевую роль в передаче нейроном нервного импульса.
Нейроны слухового нерва переключаются на нейроны продолговатого мозга – улитковые ядра. Причем улитковые ядра – последние образования слухового анализатора, получающие нервные импульсы только от одного уха.
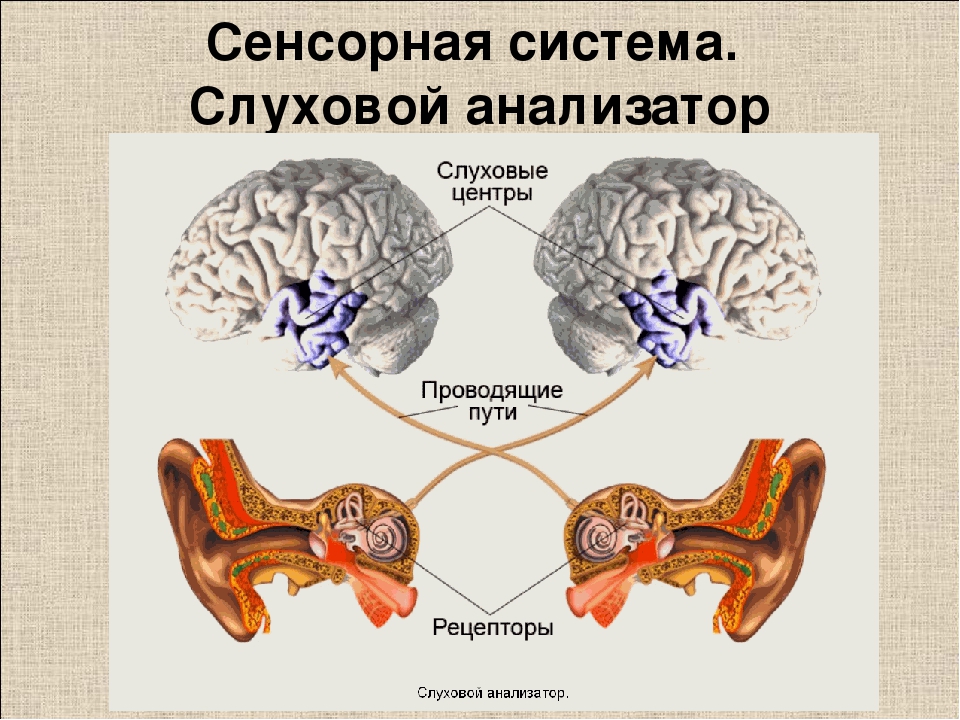
Проводящие пути и подкорковые центры слухового анализатора является частью центральной нервной системы (ЦНС) и включает восходящую (афферентную) и нисходящую (эфферентную) системы. Анатомически, он находится в стволе головного мозга, подкорковых структурах головного мозга. Упрощенная схема восходящей слуховой системы показана на схеме.
Анатомически, он находится в стволе головного мозга, подкорковых структурах головного мозга. Упрощенная схема восходящей слуховой системы показана на схеме.
Как видно из схемы, количество нервных клеток (нейронов) многократно возрастает по мере возвышения от слухового нерва до коры головного мозга. В слуховом нерве их примерно 35 тысяч, а в слуховой коре – более 12 миллионов. Кроме того, по мере возвышения к слуховой коре возрастает и связь слуховых нейронов как между обоими сторонами мозга, так и с нейронами других сенсорных систем, зонами памяти, речи и многими другими.
Примечательно, что выше правого слухового нерва и ядер улитки, в которых его нейроны переключаются на следующий уровень, основная часть восходящих слуховых нейронов переходят со стороны этого уха на левую сторону мозга. И наоборот. Таким образом, происходит «перекрест» проводящих путей слухового анализатора, что хорошо видно и из схемы ствола головного мозга.
Центральный отдел органа слуха
Центральный (корковый) отдел слухового анализатора расположен в височных долях коры головного мозга. Нервные импульсы от правого уха попадают главным образом в левое полушарие мозга, и наоборот, от левого уха – в правое. Это имеет большое значение при слухопротезировании, и вот почему. Слуховые зоны обоих полушарий выполняют хотя и аналогичную, но разную работу.
Нервные импульсы от правого уха попадают главным образом в левое полушарие мозга, и наоборот, от левого уха – в правое. Это имеет большое значение при слухопротезировании, и вот почему. Слуховые зоны обоих полушарий выполняют хотя и аналогичную, но разную работу.
Исследования 1960-70-х годов показали, что у большинства правшей левое полушарие лучше обрабатывает высокочастотные, быстро изменяющиеся звуки, и лучше воспринимает отдельные звуки, слоги и слова речи. Именно поэтому левое полушарие и соответственно правое ухо назвали доминантными по восприятию речи. И именно поэтому, у большинства правшей в случае невозможности бинаурального слухопротезирования преимущественным является слухопротезирование правого уха. У левшей – как правило наоборот. Но поскольку существует много индивидуальных различий, при аудиометрическом обследовании необходимо определить какое ухо лучше воспринимает словесные тесты. Оценка же восприятия целостной речи является достаточно долгим и непростым психоакустическим исследованием и в сурдологической практике не применяется.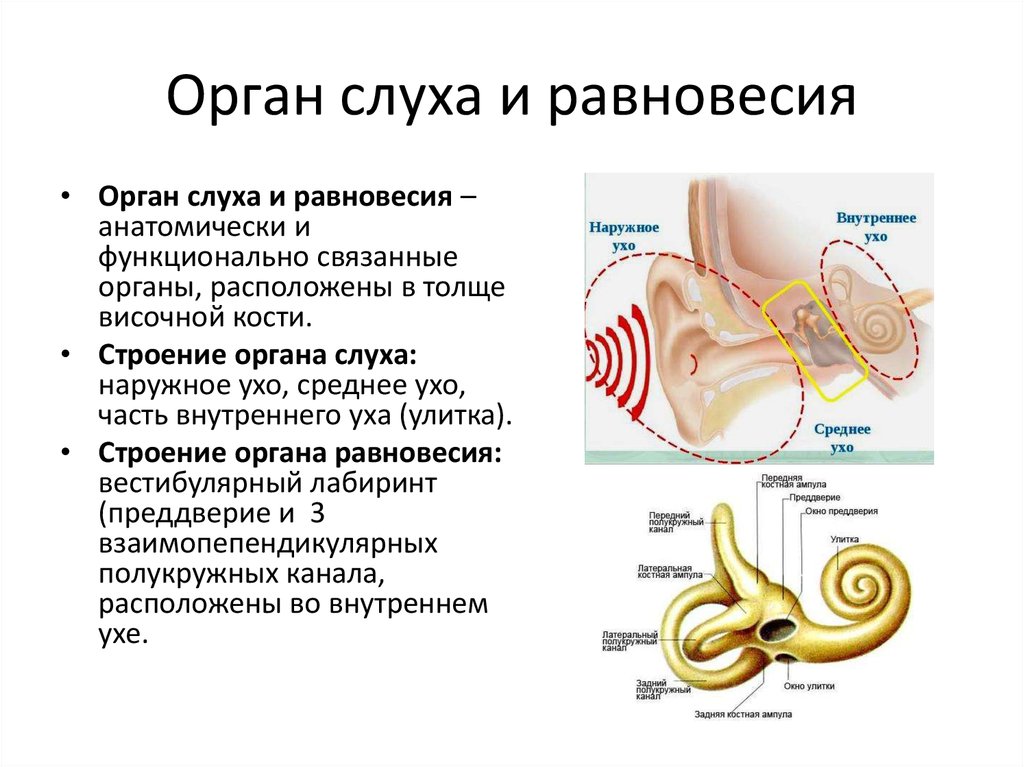
Более поздние исследования в 1970-80-е годы, показали, что с речевой доминантностью полушарий не все так просто.
Эксперименты многих ученых показали, что полушарие, противоположное доминантному при восприятии отдельных слов, (у большинства правшей – правое) гораздо лучше воспринимает интонацию, ритм речи, которые необходимы для понимания того, утверждает ли что-то говорящий или спрашивает, серьезно ли говорит или шутит. То есть оно лучше понимает предложения в целом. Более того, именно противоположное речевому доминантному полушарие связывает все предложения в общий смысл всего сказанного, например, весь рассказ, весь разговор в целом. Таким образом, считавшееся «доминантным» полушарие (левое у правшей) осуществляет последовательный анализ отдельных звуков, а считавшееся «не доминантным» – целостное восприятие речевых сообщений.
Функции уха
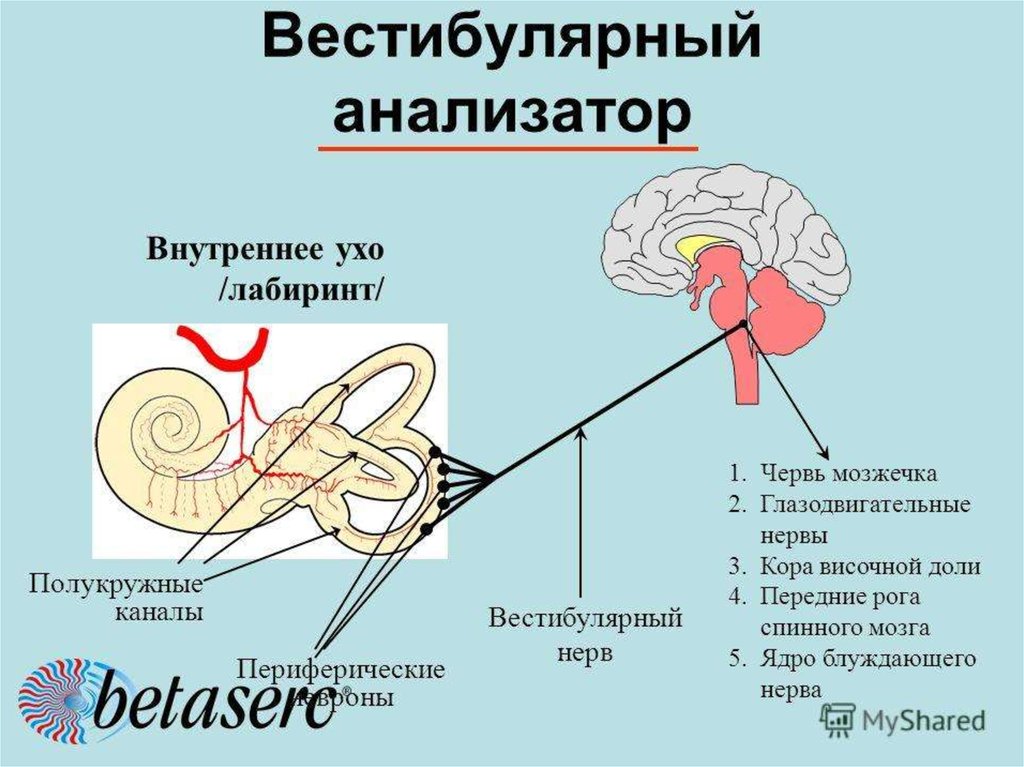
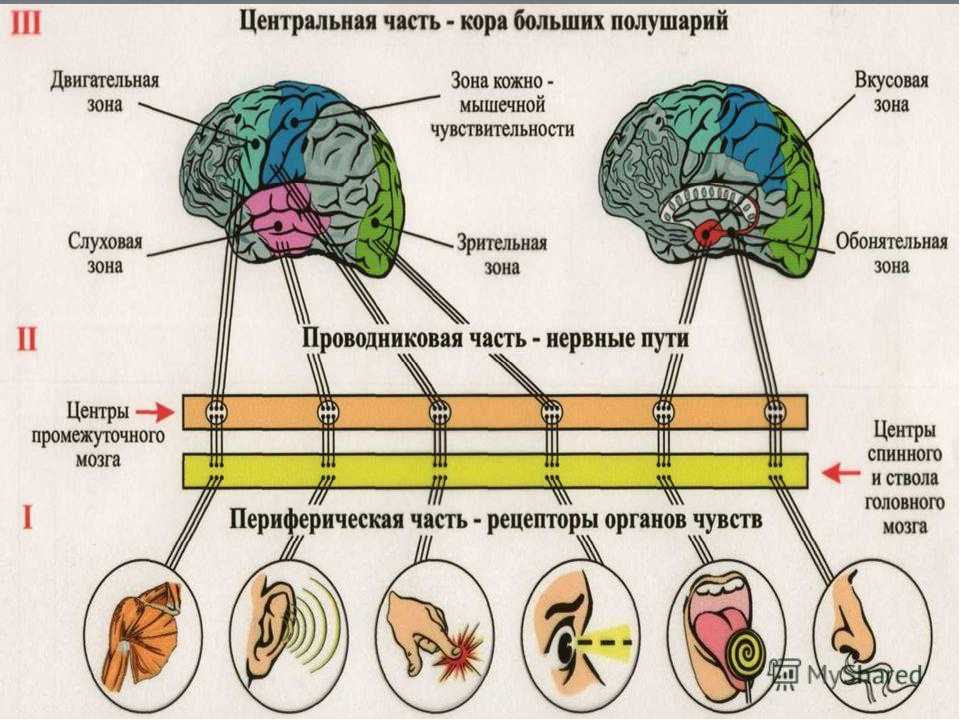
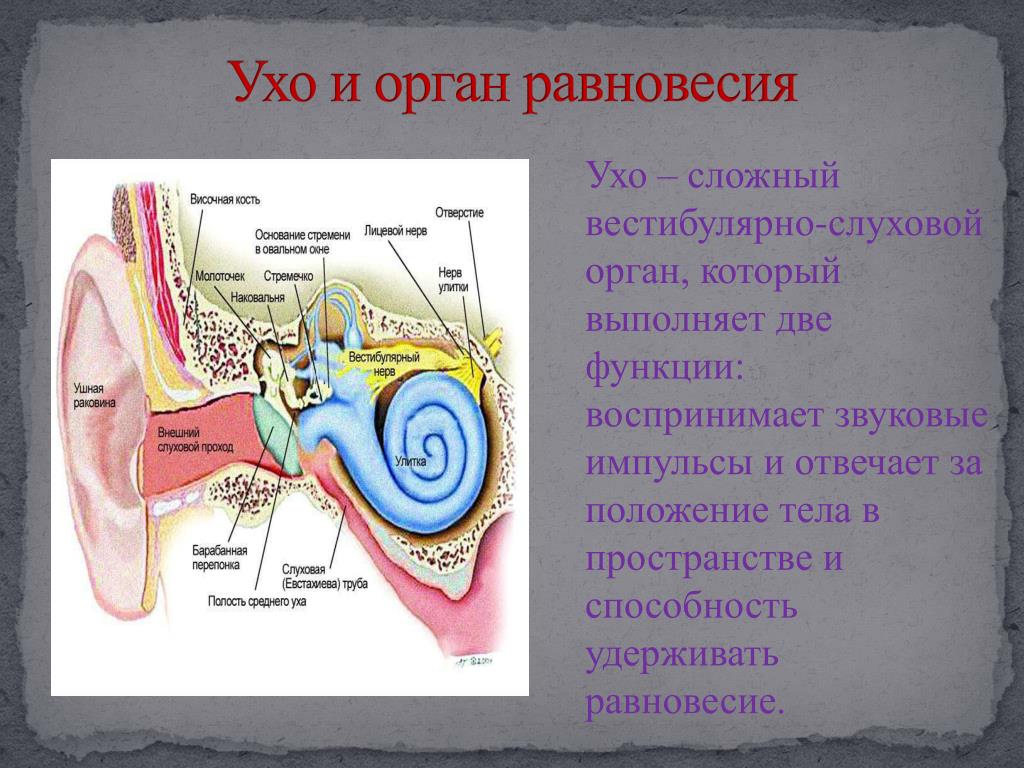
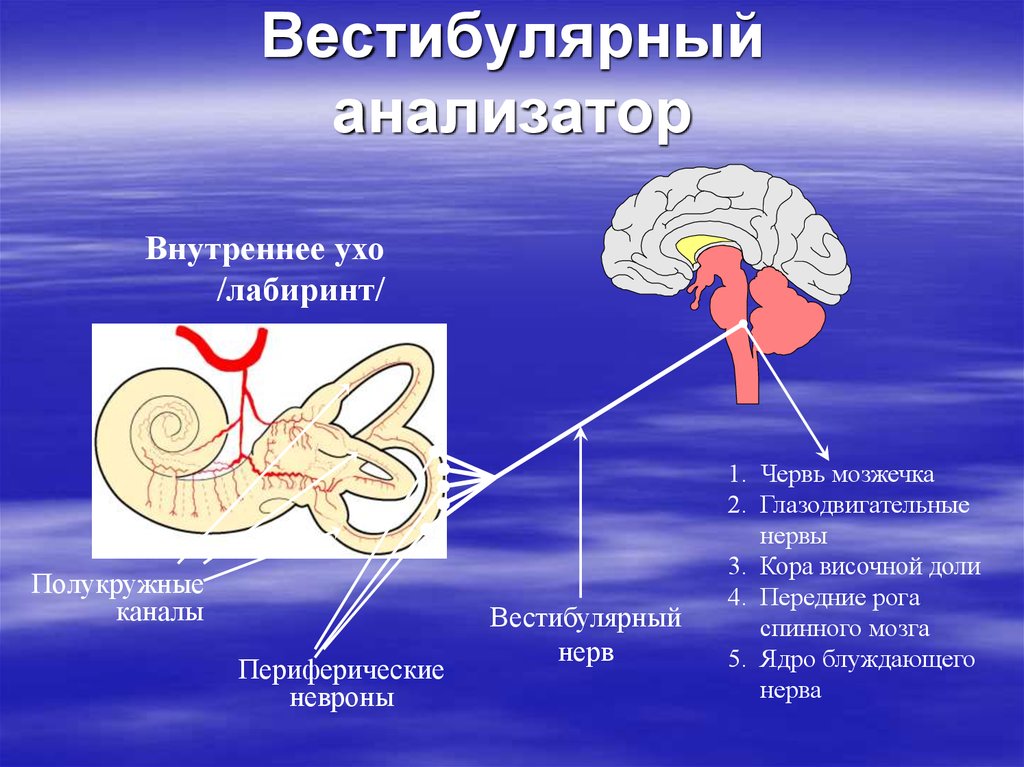
Ухо является одним из органов чувств, с помощью которого человек получает очень важную информацию из внешнего мира. Оно выполняет две функции: слуховую и вестибулярную (функцию ориентирования организма и поддержания равновесия тела в пространстве). В соответствии с этим, в ухе находятся конечные отделы двух анализаторов: слухового и вестибулярного.
Оно выполняет две функции: слуховую и вестибулярную (функцию ориентирования организма и поддержания равновесия тела в пространстве). В соответствии с этим, в ухе находятся конечные отделы двух анализаторов: слухового и вестибулярного.
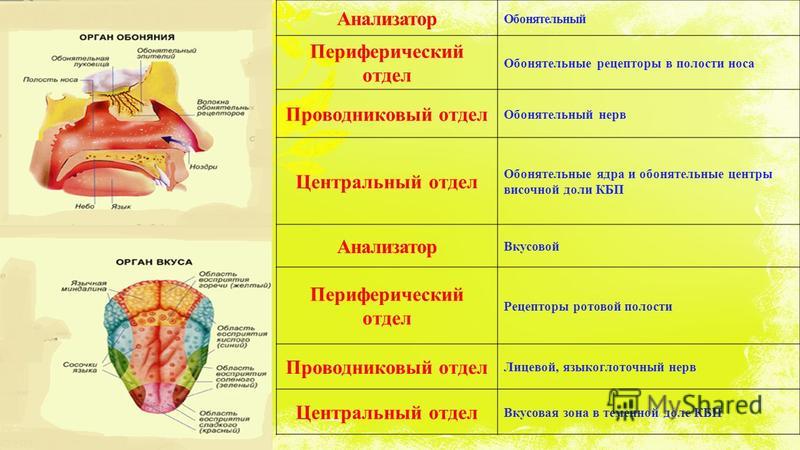
Слуховой анализатор — это анализатор, который позволяет проводить анализ сигнала на расстоянии. Что раздражает слуховой анализатор? Конечно же- звук.
Раздражение слухового анализатора имеет свои закономерности:
1. Слуховой диапазон человеческого уха. Если число колебаний окружающей среды выше 16 в секунду, то здоровый человек воспринимает эти колебания как звук. Если они превышают число 20 тыс. в секунду, то человек их слышать перестает.
2. Переменная чувствительность к звукам разной частоты. Наибольшая чувствительность органа слуха человека к звукам частотой 1000 — 3000 в секунду.
3. Человеческое ухо способно различать абсолютную высоту звука, определять интервалы и направление источника звука.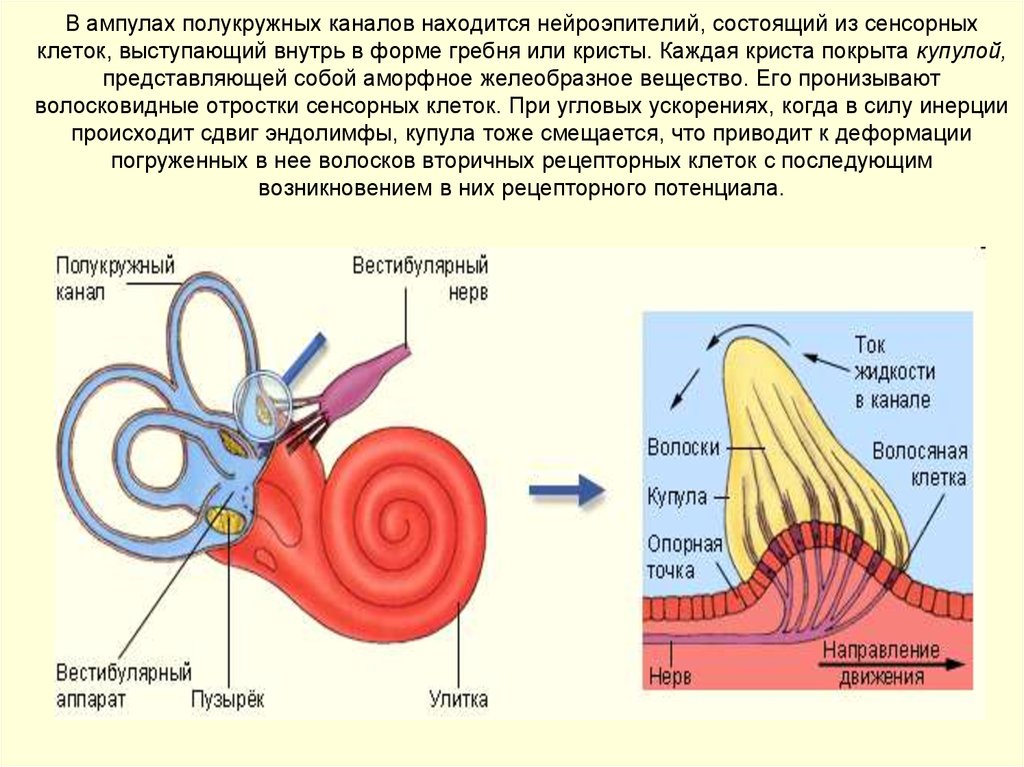
Периферический отдел, проводящий путь, корковый конец — это составляющие слухового анализатора.
В свою очередь периферический отдел делится на 2 аппарата: звукопроводящий и звуковоспринимающий.
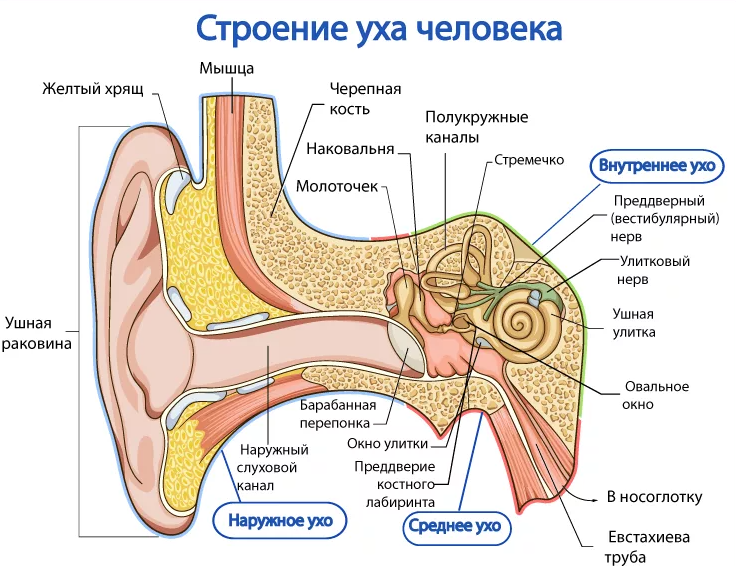
К звукопроводящему аппарату относят:
- внешнее ухо;
- среднее ухо;
- пери- и эндолимфатические пространства внутреннего уха;
- базилярную пластинку;
- присинковую мембрану улитки.
Доставка звука к рецептору — главная задача звукопроводящего аппарата. Звуковоспринимающий аппарат представлен периферическим рецепторным органом — спиральным органом. Звуковоспринимающий аппарат трансформирует механические колебания в процесс нервного возбуждения.
Теперь рассмотрим подробнее функции ушей и отдельных его частей.
Функции наружного уха:
- сбор звуков.
Ушная раковина собирает и направляет звуковые волны к слуховому проходу.
- проводящая.
Наружный слуховой канал проводит звуковые колебания к среднему уху.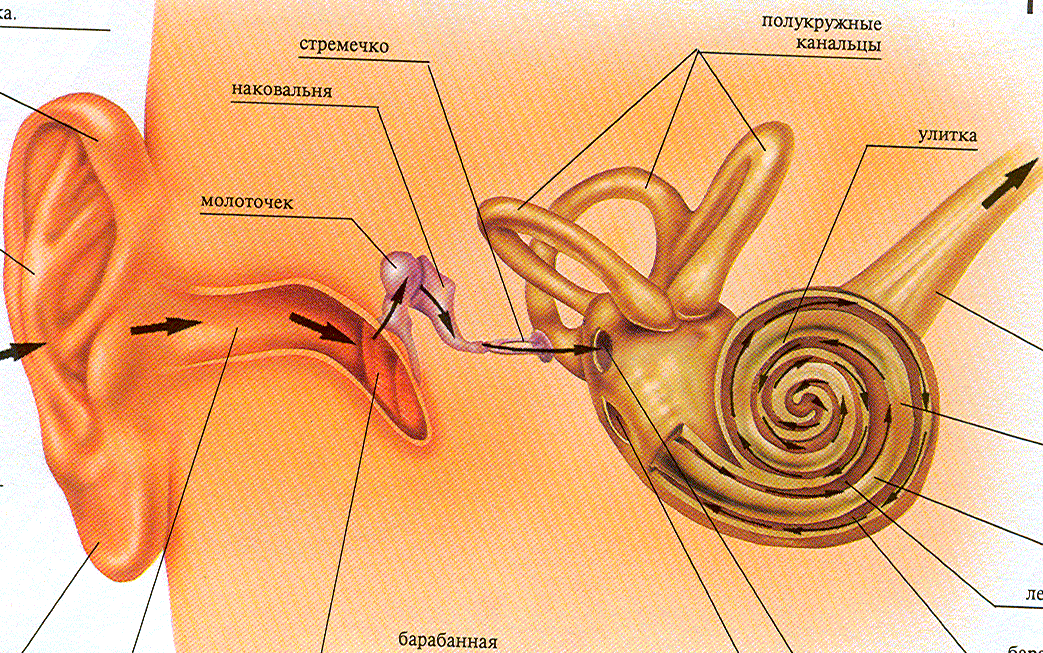
- защитная.
В наружном слуховом проходе железами производится ушная сера, которая защищает ухо от негативного внешнего воздействия и микроорганизмов.
Функции среднего уха:
Среднее ухо — это полость, которая от наружного уха отделяется барабанной перепонкой. Строение среднего уха состоит из системы слуховых косточек: молоточек, наковальня, стремя.
В среднее ухо открывается отверстие евстахиевой трубы, которая обеспечивает соединение среднего уха с окружающей средой через носоглотку. Именно это выравнивает давление давление на барабанную перепонку с двух сторон: извне и со стороны среднего уха. Отверстия евстахиевых труб открываются при глотании, жевании, зевоте. Именно поэтому на самолетах при подъеме на большую высоту советуют пить воду небольшими глотками, или сосать кислые конфеты, которые вызывают выделение слюны. При ее глотании возникает выравнивания давления, исключая болевые раздражения в области барабанных перепонок.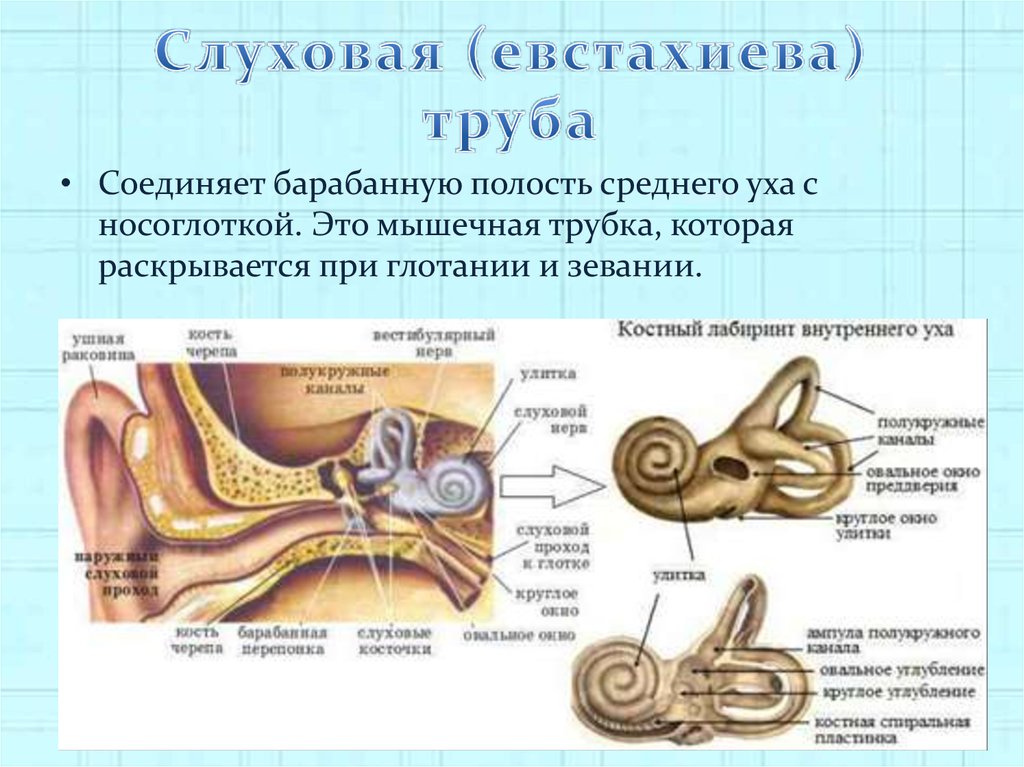
Функции евстахиевой трубы:
- вентиляционная
- дренажная (способствует оттоку жидкости из полости среднего уха)
- защитная
Функции среднего уха представляют собой по сути функции слуховых косточек:
- звукопроводящая.
Заключается в проведении звуковых колебаний от наружного уха к внутреннему.
- трансформационная.
Слуховые косточки изменяют звуковые колебания таким образом, что их амплитуда уменьшается, а интенсивность увеличивается. В целом интенсивность звука в среднем ухе увеличивается в 22 раза, или на 39,7 дБ.
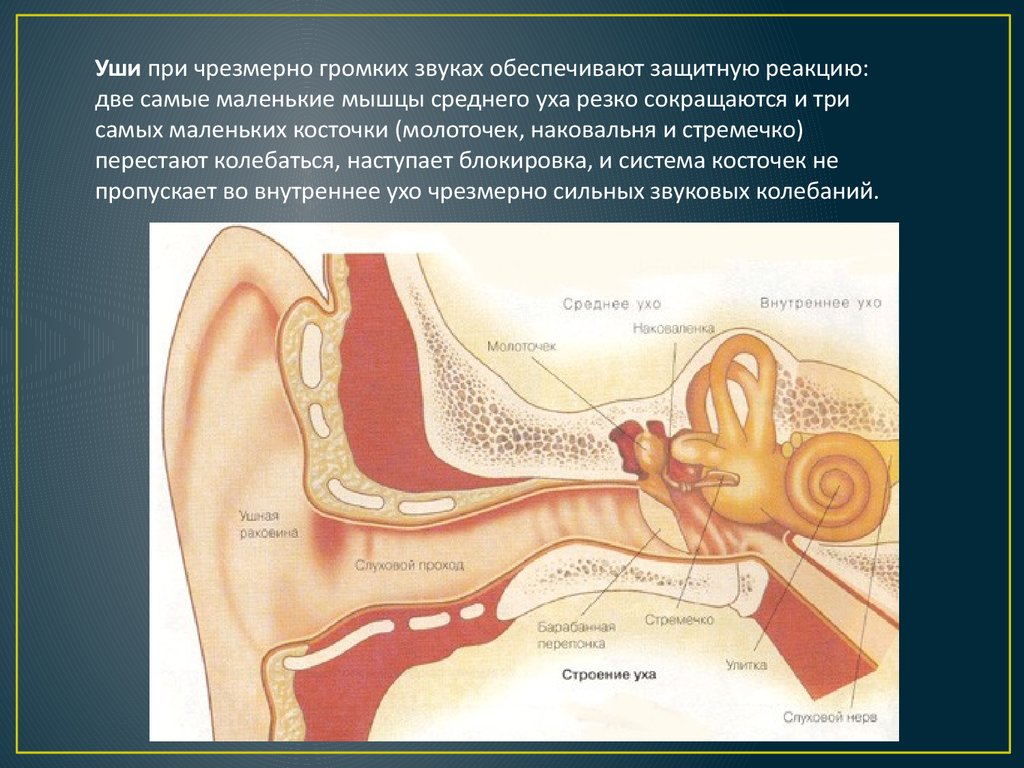
- защитная.
Эту функцию выполняют мышцы среднего уха. В случае большой интенсивности звука они рефлекторно резко сокращаются. Это приводит к торможению колебания барабанной перепонки и слуховых косточек и, соответственно, к уменьшению звукового давления, что передается к перилимфе.
Функции внутреннего уха:
Строение внутреннего уха включает в себя преддверие, улитку, полукружные каналы и слуховые нервы. Вся эта система обеспечивает передачу звуковой информации из уха к мозгу. Таким образом, основная функция внутреннего уха — звуковоспринимающая.
Существуют два пути проведения звуков в лабиринт:
- воздушная проводимость
через наружный слуховой проход, барабанную перепонку и цепь слуховых косточек
- тканевая проводимость
непосредственно через ткани черепа
Наши органы слуха играют важную роль не только в элементарном восприятии и воспроизведении звуков окружающей среды. Это еще и мощный инструмент регулирования баланса и равновесия человека.
Слуховые рецепторы
Слуховые рецепторыРеклама
1 из 15
Верхний вырезанный слайд
Скачать для чтения офлайн Реклама
Реклама
Слуховые рецепторы
- СЛУХОВЫЕ РЕЦЕПТОРЫ
• А.
 Внутреннее ухо:
• Слуховая и вестибулярная системы
тесно связаны. Рецепторы для
оба расположены в височной кости, в
извилистая камера, называемая костной
лабиринт. Деликатный непрерывный
мембрана подвешена внутри кости
лабиринт, создающий вторую камеру
внутри первого. Эта камера называется
перепончатый лабиринт. Целиком
заполненная жидкостью структура называется внутренней
ухо.
• Внутреннее ухо имеет два покрытых мембраной
выходы в заполненное воздухом среднее ухо —
овальное окно и круглое окно.
Овальное окно заполнено пластиной из
стремечко, третья косточка среднего уха.
стремечко вибрирует в ответ на вибрации
барабанной перепонки, закрепляя жидкость
внутреннее ухо хлюпает вперед и назад.
Внутреннее ухо:
• Слуховая и вестибулярная системы
тесно связаны. Рецепторы для
оба расположены в височной кости, в
извилистая камера, называемая костной
лабиринт. Деликатный непрерывный
мембрана подвешена внутри кости
лабиринт, создающий вторую камеру
внутри первого. Эта камера называется
перепончатый лабиринт. Целиком
заполненная жидкостью структура называется внутренней
ухо.
• Внутреннее ухо имеет два покрытых мембраной
выходы в заполненное воздухом среднее ухо —
овальное окно и круглое окно.
Овальное окно заполнено пластиной из
стремечко, третья косточка среднего уха.
стремечко вибрирует в ответ на вибрации
барабанной перепонки, закрепляя жидкость
внутреннее ухо хлюпает вперед и назад.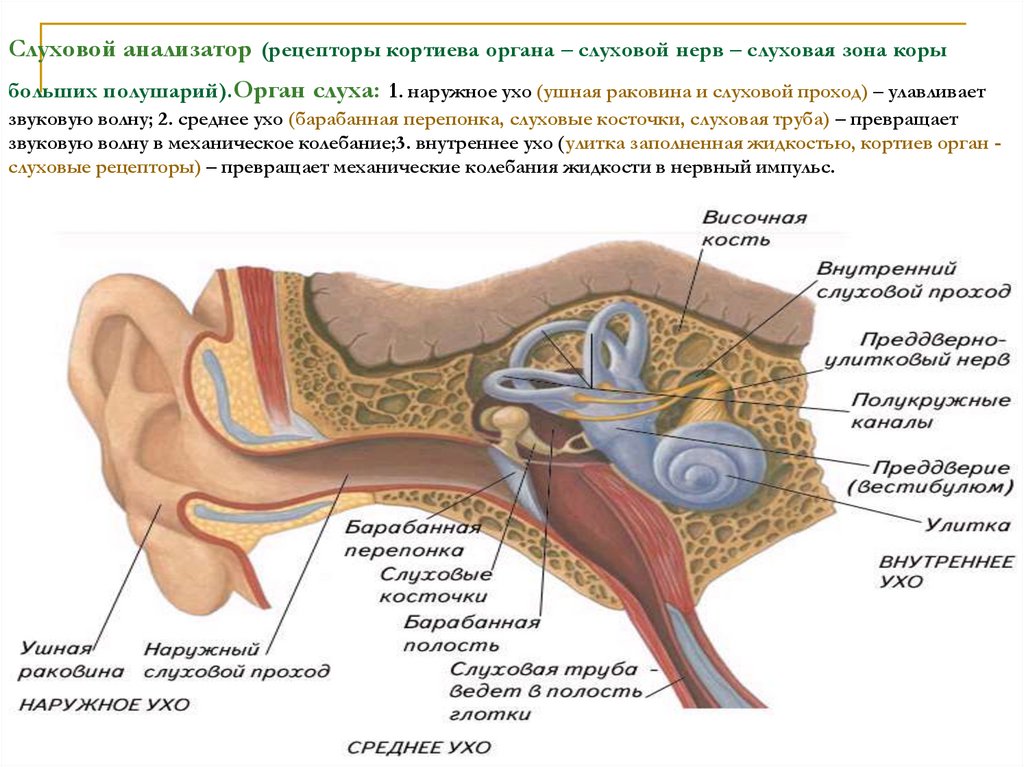
- Овальное окно открывается в большую центральную область внутреннего уха, называемую преддверием. Все
органы внутреннего уха ответвляются от этой центральной камеры. С одной стороны находится улитка, с другой
другие полукружные каналы. Прилегают маточка и мешочек, дополнительные вестибулярные органы.
в вестибюль.
Перепончатый лабиринт заполнен особой жидкостью, называемой эндолимфой. Эндолимфа очень
похож на внутриклеточную жидкость: в ней много калия и мало натрия. Ионный состав
необходим для оптимального функционирования вестибулярных и слуховых волосковых клеток. Пространство между
перепончатые и костные лабиринты заполнены перилимфой, которая очень похожа на нормальную
спинномозговая жидкость.
Б. Слуховая трансдукция:
Преобразование звука в нервный сигнал происходит в улитке.
Если бы мы развернули улитку в форме улитки, она выглядела бы так:
Когда стремечко вибрирует в овальном окне, перилимфа хлюпает вперед и назад, вызывая вибрацию
круглое окно в дополнительном ритме.
- Перепончатый лабиринт улитки окружает
заполненная эндолимфой лестничная клетка. Два отсека
костный лабиринт, в котором находится перилимфа,
называются вестибулярными и барабанными лестницами. Внутри скалы
media — рецепторный орган, кортиев орган. Он отдыхает
на части перепончатого лабиринта, базилярного
мембрана.
Один оборот улитки обведен синим цветом.
Вы можете увидеть слуховой нерв, выходящий из основания
улитка; он пройдет через височную кость к
мозговой ствол.

- Базилярная мембрана на самом деле тоньше и уже у основания улитки, чем на кончике
(вершина), которая кажется назад, учитывая, что улитка имеет наибольшую ширину у основания. Свойства
базилярная мембрана изменяется по мере изменения ее формы; как и гитарные струны, тонкие вещи вибрируют
до высоких тонов, а толстые предметы вибрируют до низких тонов. Это означает, что базилярная мембрана
вибрирует до высоких частот у основания улитки и до низких частот у верхушки. Волосы
клетка в основании улитки будет лучше всего реагировать на высокие частоты, так как на этих частотах
базилярная мембрана под ним будет вибрировать больше всего. Основная идея заключается в том, что хотя волосы
клетки располагаются по порядку вдоль базилярной мембраны, от высокочастотных к низкочастотным,
именно свойства базилярной мембраны определяют этот градиент, а не свойства
волосковые клетки.
Наша способность различать две близкие частоты на самом деле намного лучше, чем можно было бы предположить.
просто из механики базилярной мембраны.
- C. Центральные слуховые пути:
Слуховой нерв передает сигнал в ствол мозга и синапсы в кохлеарном ядре.
От кохлеарного ядра слуховая информация разделяется как минимум на два потока, подобно
зрительные пути разделены на движение и обработку формы. Волокна слухового нерва идут к
синапсы вентрального ядра улитки на клетках-мишенях с гигантскими, похожими на руки окончаниями.
Что-то в этом плотном соединении позволяет сохранять синхронизацию сигнала до
микросекунды (потенциалы действия порядка миллисекунд, так что это немалый подвиг).
клетки вентрального ядра улитки затем проецируются на совокупность ядер в мозговом веществе, называемую
превосходная олива. В верхней оливе мельчайшие различия во времени и громкости
звук в каждом ухе сравнивается, и по этому можно определить направление звука
от.
- Обратите внимание, что оба пути двусторонние. Следствие
заключается в том, что поражения в любом месте пути обычно
не оказывают заметного влияния на слух. Глухота по существу
только при поражении среднего уха, улитки или
слуховой нерв.
Оба потока информации от нижнего холмика
перейти к сенсорному таламусу. Слуховое ядро
таламус — медиальное коленчатое тело. Медиальный
проекты коленчатого тела в первичную слуховую кору, расположенную на
Имейте в виду, когда вы пытаетесь вспомнить этот путь, что все слуховые ядра, по-видимому, имеют аналоги в
банки височных долей.
 Каждый канал устроен как
показан ниже в виде сплошного кольца, заполненного эндолимфой. Настоящие волосковые клетки располагаются в небольшом вздутии у основания.
называется ампулой.
2. ВОР
Хотя VOR воздействует на все три пары мышц, пара медиально-латеральных прямых мышц, соединенная с горизонтальной
канал, геометрически проще всего нарисовать
Каждый канал устроен как
показан ниже в виде сплошного кольца, заполненного эндолимфой. Настоящие волосковые клетки располагаются в небольшом вздутии у основания.
называется ампулой.
2. ВОР
Хотя VOR воздействует на все три пары мышц, пара медиально-латеральных прямых мышц, соединенная с горизонтальной
канал, геометрически проще всего нарисовать - Наружное ухо
Звуковая энергия распространяется от своих источников. Для точечного источника звука он распространяется согласно
закон обратных квадратов. При заданной интенсивности звука большее ухо улавливает большую часть волны и, следовательно, больше звуковой энергии.
Структуры внешнего уха действуют как часть предварительного усилителя уха, повышая чувствительность слуха.
Слуховой проход действует как замкнутый трубчатый резонатор, усиливая звуки в диапазоне 2-5
килогерц.
Барабанная перепонка
Барабанная перепонка или «барабанная перепонка» получает
колебания, распространяющиеся по слуховому проходу и
переносит их через крошечные косточки в
овальное окно, порт во внутреннее ухо.
 барабанная перепонка примерно в пятнадцать раз больше, чем
овальное окно, дающее усиление около
пятнадцать по сравнению с одним только овальным окном.
барабанная перепонка примерно в пятнадцать раз больше, чем
овальное окно, дающее усиление около
пятнадцать по сравнению с одним только овальным окном. - Резонанс слухового канала
Области максимальной чувствительности человеческого слуха можно смоделировать в виде закрытой трубки.
резонанс слухового прохода. Наблюдаемый пик около 3700 Гц на теле
температура соответствует трубке длиной 2,4 см. Более высокая частота
пик чувствительности находится на частоте около 13 кГц, что несколько выше расчетной 3-й частоты.
гармоника закрытого цилиндра
косточки
Три мельчайшие кости в теле образуют связь между вибрацией
барабанная перепонка и силы, действующие на овальное окно внутреннего уха.
С помощью достаточно длинного рычага вы можете поднять большой камень с небольшим усилием, приложенным к рычагу.
другой конец рычага.
 Усиление силы можно изменить, сместив шарнир
точка.
Косточки можно рассматривать как составной рычаг, который обеспечивает умножение
сила. Считается, что это действие рычага обеспечивает усиление примерно в
три в оптимальных условиях, но может быть скорректировано мышечным действием до фактического
ослабить звуковой сигнал для защиты от громких звуков.
Усиление силы можно изменить, сместив шарнир
точка.
Косточки можно рассматривать как составной рычаг, который обеспечивает умножение
сила. Считается, что это действие рычага обеспечивает усиление примерно в
три в оптимальных условиях, но может быть скорректировано мышечным действием до фактического
ослабить звуковой сигнал для защиты от громких звуков. - Вибрация косточек Вибрация барабанной перепонки является передается в овальное окно внутреннее ухо с помощью косточек, которые достигают усиления за счет действие рычага. Рычаг регулируется под действием мышц и может фактически приглушать громкие звуки для защиты ухо. Внутреннее ухо
- Внутреннее ухо можно представить как два
органы: полукружные каналы, служащие
как орган равновесия тела и улитка
который служит микрофоном тела,
преобразование импульсов звукового давления из
наружное ухо в электрические импульсы, которые
передаются в мозг через
слуховой нерв.
 Базилярная мембрана внутреннего уха играет
решающую роль в восприятии высоты тона
по теории места.
Полукружные каналы
Полукружные каналы – это
органы равновесия, обнаруживающие ускорение в
трех перпендикулярных плоскостях. Эти
акселерометры используют волосковые клетки
аналогичны таковым на кортиевом органе, но
эти волосковые клетки обнаруживают движения
жидкость в каналах, вызванная
угловое ускорение вокруг оси
перпендикулярно плоскости канала.
Крошечные плавающие частицы помогают процессу
стимулируя волосковые клетки, когда они двигаются
жидкость. Каналы соединены с
слуховой нерв.
Базилярная мембрана внутреннего уха играет
решающую роль в восприятии высоты тона
по теории места.
Полукружные каналы
Полукружные каналы – это
органы равновесия, обнаруживающие ускорение в
трех перпендикулярных плоскостях. Эти
акселерометры используют волосковые клетки
аналогичны таковым на кортиевом органе, но
эти волосковые клетки обнаруживают движения
жидкость в каналах, вызванная
угловое ускорение вокруг оси
перпендикулярно плоскости канала.
Крошечные плавающие частицы помогают процессу
стимулируя волосковые клетки, когда они двигаются
жидкость. Каналы соединены с
слуховой нерв. - Сонарное обнаружение у летучих мышей НАЧАЛО. Расстояние между импульсами давления у летучей мыши ультразвуковые щебеты и эхо (представленные белой и красной волной очертания соответственно) определяет, насколько мала цель, которую хищник может обнаружить.
- Летучие мыши ориентируются с помощью отраженных звуковых волн. Этот процесс, известный как эхолокация,
позволяет этим животным «видеть» в темноте.
 Чтобы обнаружить предметы, летучие мыши должны сначала испустить
серия звуковых импульсов. Эти импульсы распространяются наружу и ударяют по объектам. Импульсы
затем отражаются от объектов и возвращаются обратно к летучим мышам. Обнаружены их большие
ушах, звуки быстро анализируются эхолокационным центром мозга. Этот
анализ настолько точен, что летучая мышь может определить местонахождение движущейся рыбы посредством критического анализа
рябь, образующаяся на поверхности воды.
Летающий слепой
Млекопитающие могут слышать более высокие частоты, чем другие существа, из-за характерного устройства
крошечных костей в ушах, а также структура их улитки или внутреннего уха. Даже среди млекопитающих
однако слуховые способности сильно различаются. Среди людей большинство молодых людей могут различать звуки с
частоты от 20 герц, или циклов в секунду, до 20 килогерц (кГц). Но даже этот самый высокий тон
низким по стандартам эхолокации: типичное щебетание гидролокатора летучей мыши использует частоты до 120 кГц, и
крики афалин включают частоты до 150 кГц или около того.
Чтобы обнаружить предметы, летучие мыши должны сначала испустить
серия звуковых импульсов. Эти импульсы распространяются наружу и ударяют по объектам. Импульсы
затем отражаются от объектов и возвращаются обратно к летучим мышам. Обнаружены их большие
ушах, звуки быстро анализируются эхолокационным центром мозга. Этот
анализ настолько точен, что летучая мышь может определить местонахождение движущейся рыбы посредством критического анализа
рябь, образующаяся на поверхности воды.
Летающий слепой
Млекопитающие могут слышать более высокие частоты, чем другие существа, из-за характерного устройства
крошечных костей в ушах, а также структура их улитки или внутреннего уха. Даже среди млекопитающих
однако слуховые способности сильно различаются. Среди людей большинство молодых людей могут различать звуки с
частоты от 20 герц, или циклов в секунду, до 20 килогерц (кГц). Но даже этот самый высокий тон
низким по стандартам эхолокации: типичное щебетание гидролокатора летучей мыши использует частоты до 120 кГц, и
крики афалин включают частоты до 150 кГц или около того.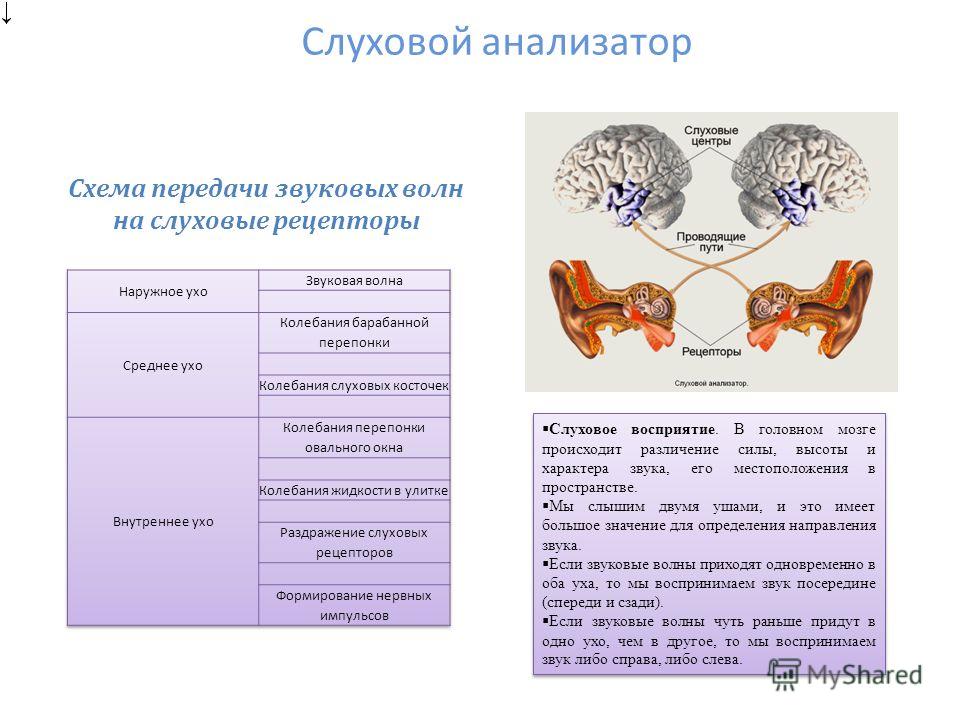 Однако птицы не могут
производить или слышать высокие коротковолновые звуки, необходимые для отслеживания целей размером с насекомое. Несколько
птицы, которые могут эхолокировать, используют более низкие частоты, и они делают это только для навигации в темноте, говорит Дж. Джордан.
Прайс, биолог из Колледжа Святой Марии в Мэриленде в городе Сент-Мэри. Даже эта ограниченная способность
однако дает преимущество, поскольку позволяет членам этих видов гнездиться в пещерах и других местах.
которые труднодоступны для хищников. За одним исключением, все птицы, способные эхолокировать, являются стрижами.
Птицы этой группы ловят насекомых на лету так же, как и летучие мыши, но делают это днем и выслеживают их.
добыча с виду, говорит Прайс. Ученые обычно полагались на характеристики помимо размера, формы и цвета, чтобы
отличить представителей одного вида стрижей от представителей другого просто потому, что у птиц
мало отличительных черт, отмечает он. До недавнего времени все стрижи, способные эхолокировать, попадали в
род Aerodramus.
Однако птицы не могут
производить или слышать высокие коротковолновые звуки, необходимые для отслеживания целей размером с насекомое. Несколько
птицы, которые могут эхолокировать, используют более низкие частоты, и они делают это только для навигации в темноте, говорит Дж. Джордан.
Прайс, биолог из Колледжа Святой Марии в Мэриленде в городе Сент-Мэри. Даже эта ограниченная способность
однако дает преимущество, поскольку позволяет членам этих видов гнездиться в пещерах и других местах.
которые труднодоступны для хищников. За одним исключением, все птицы, способные эхолокировать, являются стрижами.
Птицы этой группы ловят насекомых на лету так же, как и летучие мыши, но делают это днем и выслеживают их.
добыча с виду, говорит Прайс. Ученые обычно полагались на характеристики помимо размера, формы и цвета, чтобы
отличить представителей одного вида стрижей от представителей другого просто потому, что у птиц
мало отличительных черт, отмечает он. До недавнего времени все стрижи, способные эхолокировать, попадали в
род Aerodramus.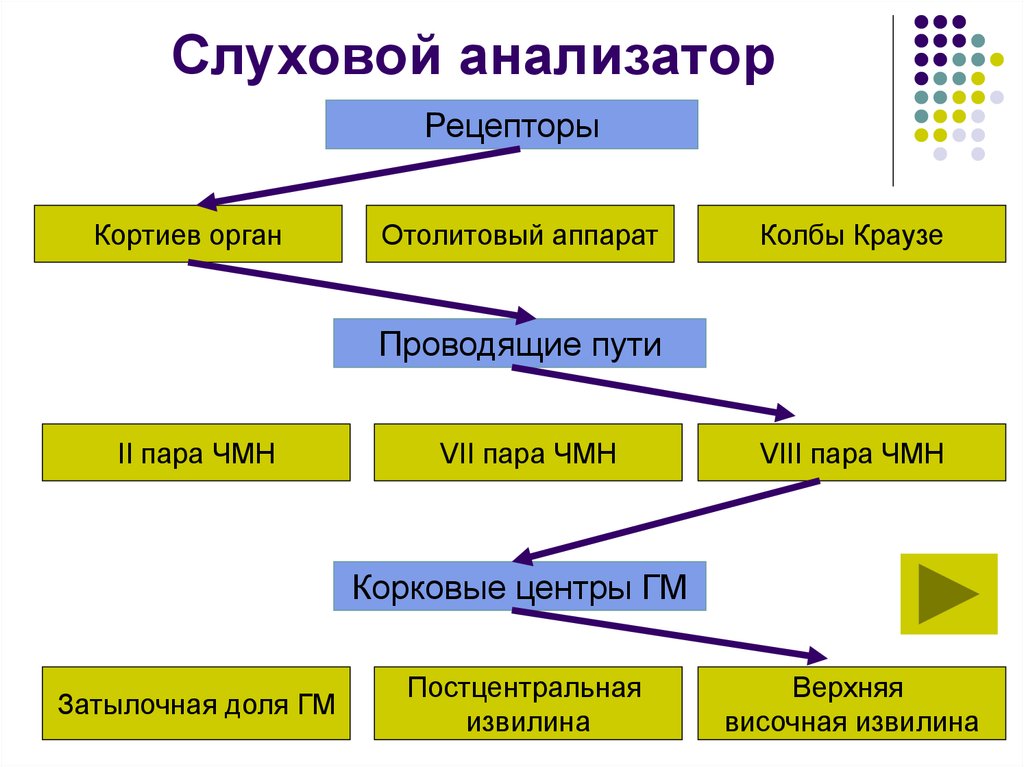 Затем Прайс и его коллеги нашли саланжа в другом роде — карликового салангана,
Collocalia troglodytes — сидит на своем гнезде в полной темноте примерно в 30 м в пещере на острове в
Филиппины.
Затем Прайс и его коллеги нашли саланжа в другом роде — карликового салангана,
Collocalia troglodytes — сидит на своем гнезде в полной темноте примерно в 30 м в пещере на острове в
Филиппины. - Эволюция глаза
- Эволюция глаза:
Когда скептики эволюции хотят напасть на теорию Дарвина, они часто указывают на человеческий глаз. Как мог
что-то настолько сложное, утверждают они, развилось в результате случайных мутаций и естественного отбора,
даже через миллионы лет?
Если эволюция происходит через градации, говорят критики, то как она могла создать отдельные части
глаз — хрусталик, сетчатка, зрачок и т. д., — поскольку ни одна из этих структур сама по себе
сделало бы зрение возможным? Другими словами, какая польза от пяти процентов глаза?
Дарвин с самого начала признавал, что его новой теории будет трудно объяснить глаз.
Трудно, но не невозможно. Ученые придумали сценарии, благодаря которым первые глазоподобные
структура, светочувствительное пигментированное пятно на коже, могло претерпеть изменения и
сложности формирования человеческого глаза с его многочисленными частями и поразительными способностями.
 В результате естественного отбора в ходе эволюции появились различные типы глаз.
человеческий глаз даже не самый лучший, с некоторых точек зрения. Поскольку кровеносные сосуды проходят через
поверхности сетчатки, а не под ней, сосуды легко разрастаются или протекают и повреждаются
зрение. Итак, говорят теоретики эволюции, антиэволюционный аргумент, что жизнь была создана
«разумный дизайнер» не выдерживает никакой критики: если бы Бог или какая-то другая всемогущая сила были ответственны за
человеческий глаз, это был какой-то неудачный дизайн.
Биологи используют ряд менее сложных светочувствительных структур, существующих сегодня у живых видов, чтобы
выдвинуть гипотезу о различных эволюционных стадиях, которые могли пройти глаза.
В результате естественного отбора в ходе эволюции появились различные типы глаз.
человеческий глаз даже не самый лучший, с некоторых точек зрения. Поскольку кровеносные сосуды проходят через
поверхности сетчатки, а не под ней, сосуды легко разрастаются или протекают и повреждаются
зрение. Итак, говорят теоретики эволюции, антиэволюционный аргумент, что жизнь была создана
«разумный дизайнер» не выдерживает никакой критики: если бы Бог или какая-то другая всемогущая сила были ответственны за
человеческий глаз, это был какой-то неудачный дизайн.
Биологи используют ряд менее сложных светочувствительных структур, существующих сегодня у живых видов, чтобы
выдвинуть гипотезу о различных эволюционных стадиях, которые могли пройти глаза. - Вот как, по мнению некоторых ученых, могли развиться некоторые глаза: простое светочувствительное пятно на
кожа какого-то древнего существа давала ему небольшое преимущество в выживании, возможно, позволяя уклоняться от
хищник. Затем случайные изменения создали углубление в светочувствительном пятне, углубляющуюся яму, которая
сделал «зрение» немного острее.
 В то же время отверстие ямы постепенно сужалось, и свет проникал внутрь.
через небольшую апертуру, как у камеры-обскуры.
Каждое изменение должно было давать преимущество в выживании, каким бы незначительным оно ни было. В конце концов, светочувствительный
пятно превратилось в сетчатку, слой клеток и пигмента в задней части человеческого глаза. Со временем линза
формируется перед глазами. Она могла возникнуть как двухслойная прозрачная ткань, содержащая
увеличивающееся количество жидкости, что придавало ему выпуклую кривизну человеческого глаза.
Фактически глаза, соответствующие каждой стадии этой последовательности, были обнаружены у существующих живых видов.
Существование этого диапазона менее сложных светочувствительных структур подтверждает гипотезу ученых.
о том, как сложные глаза, подобные нашим, могли развиваться. Первые животные с чем-то, напоминающим глаз, жили
около 550 миллионов лет назад. И, по подсчетам одного ученого, всего 364 000 лет.
были необходимы для того, чтобы похожий на камеру глаз развился из светочувствительного пятна.
В то же время отверстие ямы постепенно сужалось, и свет проникал внутрь.
через небольшую апертуру, как у камеры-обскуры.
Каждое изменение должно было давать преимущество в выживании, каким бы незначительным оно ни было. В конце концов, светочувствительный
пятно превратилось в сетчатку, слой клеток и пигмента в задней части человеческого глаза. Со временем линза
формируется перед глазами. Она могла возникнуть как двухслойная прозрачная ткань, содержащая
увеличивающееся количество жидкости, что придавало ему выпуклую кривизну человеческого глаза.
Фактически глаза, соответствующие каждой стадии этой последовательности, были обнаружены у существующих живых видов.
Существование этого диапазона менее сложных светочувствительных структур подтверждает гипотезу ученых.
о том, как сложные глаза, подобные нашим, могли развиваться. Первые животные с чем-то, напоминающим глаз, жили
около 550 миллионов лет назад. И, по подсчетам одного ученого, всего 364 000 лет.
были необходимы для того, чтобы похожий на камеру глаз развился из светочувствительного пятна.
Реклама
Слух и равновесие | Ключ Basicmedical
НАРУЖНОЕ И СРЕДНЕЕ УХО
Наружное ухо направляет звуковые волны в наружный слуховой проход (Рисунок 10–1) . У некоторых животных уши могут двигаться, как антенны радара, чтобы искать звук. От наружного слухового прохода звуковые волны проходят внутрь к барабанной перепонке (барабанная перепонка).
РИСУНОК 10–1
Структуры наружной, средней и внутренней частей человеческого уха. Звуковые волны распространяются от наружного уха к барабанной перепонке через наружный слуховой проход. Среднее ухо представляет собой заполненную воздухом полость в височной кости; содержит слуховые косточки. Внутреннее ухо состоит из костного и перепончатого лабиринтов. Чтобы прояснить взаимосвязь, улитка была немного повернута, а мышцы среднего уха опущены. (Воспроизведено с разрешения Fox SI. Физиология человека .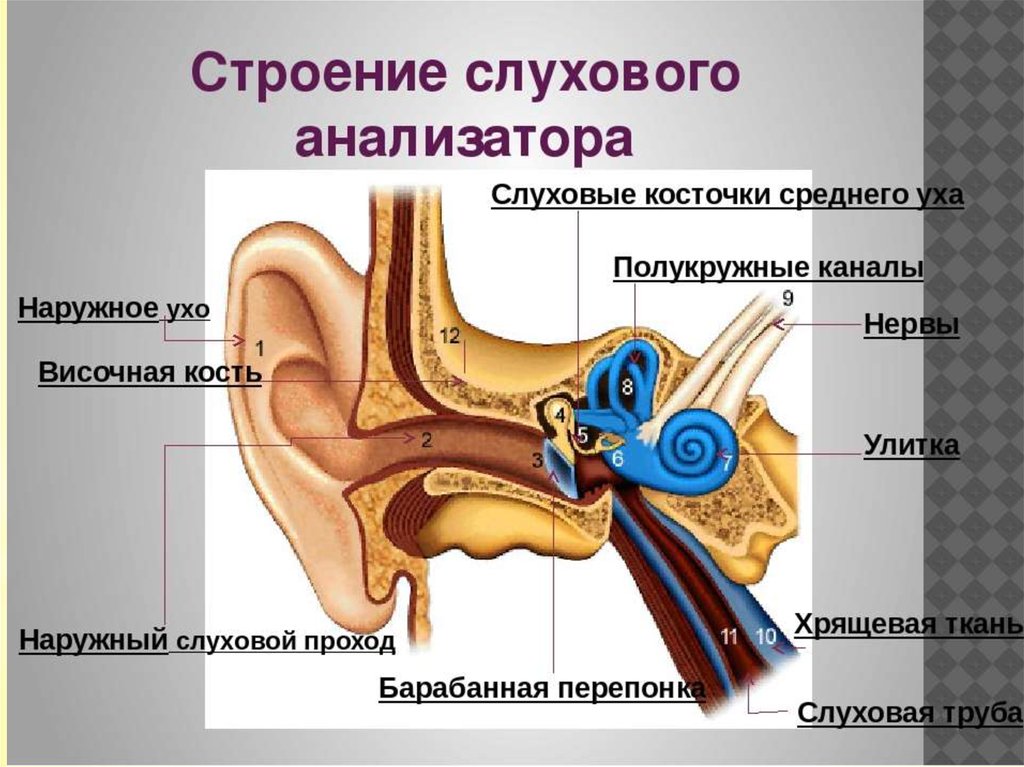 Нью-Йорк, штат Нью-Йорк: McGraw-Hill; 2008.)
Нью-Йорк, штат Нью-Йорк: McGraw-Hill; 2008.)
Среднее ухо представляет собой заполненную воздухом полость в височной кости, которая открывается через евстахиеву (слуховую) трубу в носоглотку и через носоглотку наружу. Трубка обычно закрыта, но при глотании, жевании и зевоте она открывается, сохраняя выравнивание давления воздуха по обеим сторонам барабанной перепонки. Три слуховых косточки, молоточек , наковальня, и стремя, расположены в среднем ухе (рис. 10–2) . Рукоятка (рукоятка молоточка) прикрепляется к задней части барабанной перепонки. Его головка прикрепляется к стенке среднего уха, а короткий отросток прикрепляется к наковальне, которая, в свою очередь, сочленяется с головкой стремени. Стремя названо так из-за сходства со стременем. Его подножка прикреплена кольцевой связкой к стенкам овального окна . Две небольшие скелетные мышцы, , напрягающая барабанную перепонку , и стременная, , , также расположены в среднем ухе.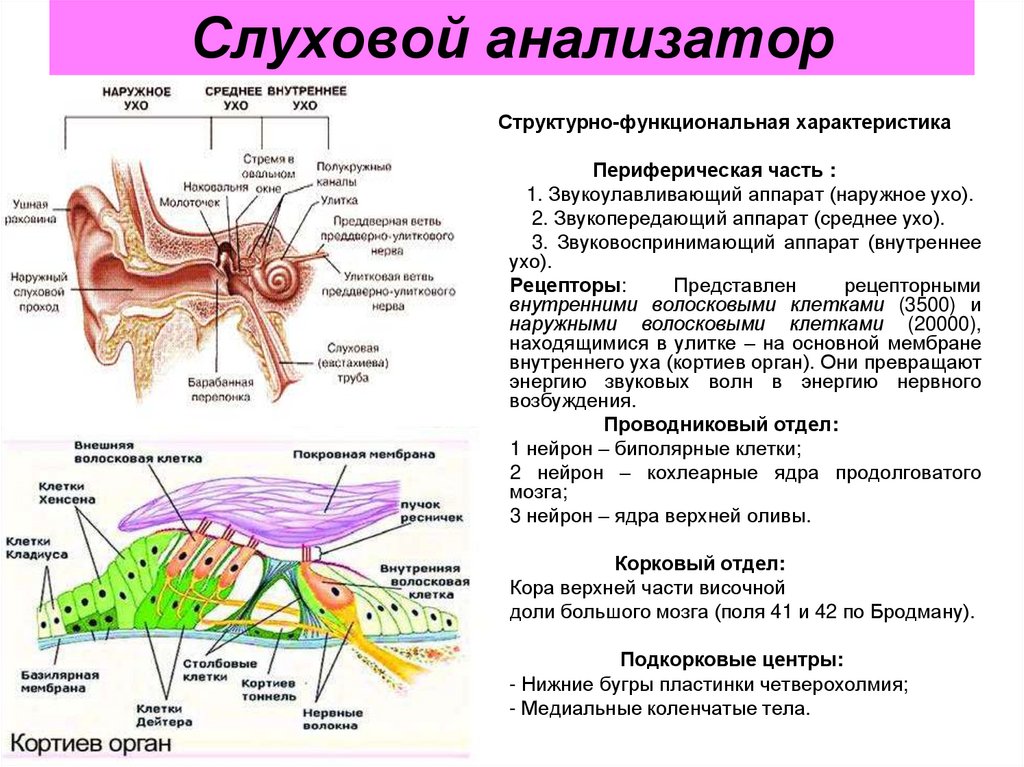 Сокращение первого тянет рукоятку молоточка медиально и уменьшает колебания барабанной перепонки; сокращение последнего вытягивает подножку стремени из овального окна. Функции косточек и мышц более подробно рассматриваются ниже.
Сокращение первого тянет рукоятку молоточка медиально и уменьшает колебания барабанной перепонки; сокращение последнего вытягивает подножку стремени из овального окна. Функции косточек и мышц более подробно рассматриваются ниже.
РИСУНОК 10–2
Среднее ухо, содержащее три слуховые косточки (молоточек, наковальню и стремечко) и две небольшие скелетные мышцы (напрягающую барабанную перепонку и стременную). Рукоятка молоточка прикрепляется к задней части барабанной перепонки. Его головка прикрепляется к стенке среднего уха, а короткий отросток прикрепляется к наковальне, которая, в свою очередь, сочленяется с головкой стремени. Подошвенная пластинка стремени кольцевидной связкой прикрепляется к стенкам овального окна. Сокращение мышцы, напрягающей барабанную перепонку, тянет рукоятку медиально и уменьшает колебания барабанной перепонки; сокращение стременной мышцы вытягивает подножку стремени из овального окна. (Воспроизведено с разрешения Fox SI.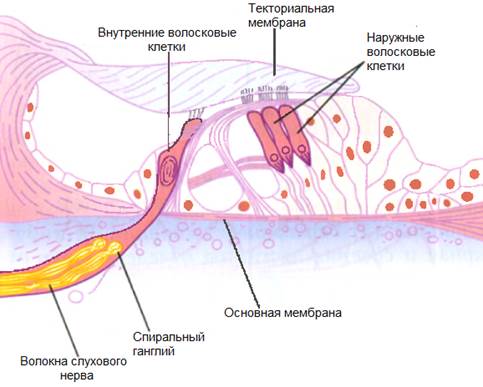 Физиология человека . Нью-Йорк, штат Нью-Йорк: McGraw-Hill; 2008.)
Физиология человека . Нью-Йорк, штат Нью-Йорк: McGraw-Hill; 2008.)
ВНУТРЕННЕЕ УХО
Внутреннее ухо (лабиринт) состоит из двух частей, одна внутри другой. Костный лабиринт представляет собой серию каналов в каменистой части височной кости и заполнен жидкостью, называемой перилимфой, , которая имеет относительно низкую концентрацию K + , сходную с таковой в плазме или спинномозговая жидкость. Внутри этих костных каналов, окруженных перилимфой, находится перепончатый лабиринт. Перепончатый лабиринт более или менее повторяет форму костных каналов и заполнен богатой K + жидкостью, называемой эндолимфой. Лабиринт состоит из трех компонентов: улитка , содержащая волосковые клетки (рецепторы) для слуха, полукружные каналы , содержащие волосковые клетки, реагирующие на вращение головы, и отолитовые органы , содержащие волосковые клетки, реагирующие на изменения в гравитация и наклон головы (Рисунок 10–3) .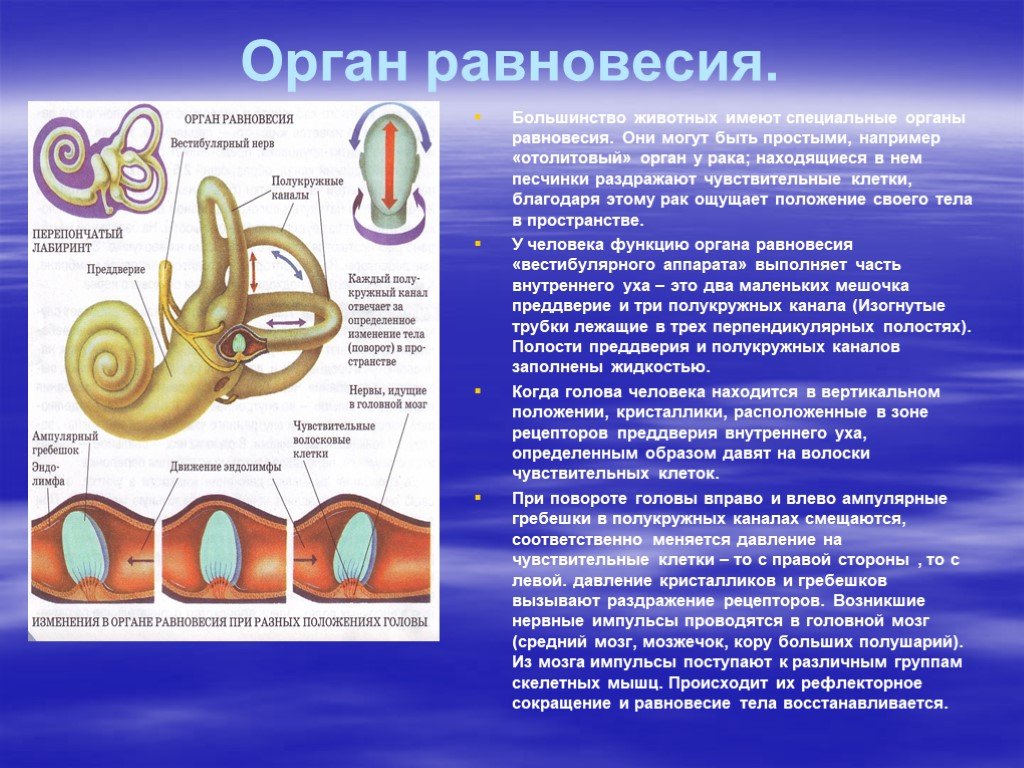
РИСУНОК 10–3
Перепончатый лабиринт внутреннего уха состоит из трех компонентов: полукружных каналов, улитки и отолитовых органов. Полукружные каналы чувствительны к угловым ускорениям, которые отклоняют студенистую купулу и связанные с ней волосковые клетки. В улитке волосковые клетки закручиваются вдоль базилярной мембраны внутри кортиева органа. Воздушные звуки приводят в движение барабанную перепонку, которая передается в улитку костями среднего уха. Это изгибает мембрану вверх и вниз. Волосковые клетки кортиева органа возбуждаются сдвигом. Отолитовые органы (мешочек и маточка) чувствительны к линейным ускорениям в вертикальной и горизонтальной плоскостях. Волосковые клетки прикреплены к отолитовой мембране. Информация от волосковых клеток улитки переносится улитковым отделом слухового (VIII черепного) нерва. Информация от волосковых клеток полукружных каналов и отолитовых органов переносится вестибулярными отделами слухового нерва.
Улитка представляет собой спиральную трубку, которая у человека имеет длину 35 мм и делает две и три четверти оборота. Базилярная мембрана и мембрана Рейсснера делят его на три камеры или лестницы (рис. 10-4) . Верхняя вестибулярная лестница и нижняя барабанная лестница содержат перилимфу и сообщаются друг с другом на вершине улитки через небольшое отверстие, называемое геликотремой . В основании улитки вестибулярная лестница заканчивается овальным окном, которое закрыто подножной пластинкой стремени. Барабанная лестница заканчивается на 9.0063 круглое окно, отверстие на медиальной стенке среднего уха, закрытое гибкой вторичной барабанной перепонкой. Медиа лестницы , средняя камера улитки, является продолжением перепончатого лабиринта и не сообщается с двумя другими лестницами.
РИСУНОК 10–4
Схема улитки и кортиева органа в перепончатом лабиринте внутреннего уха.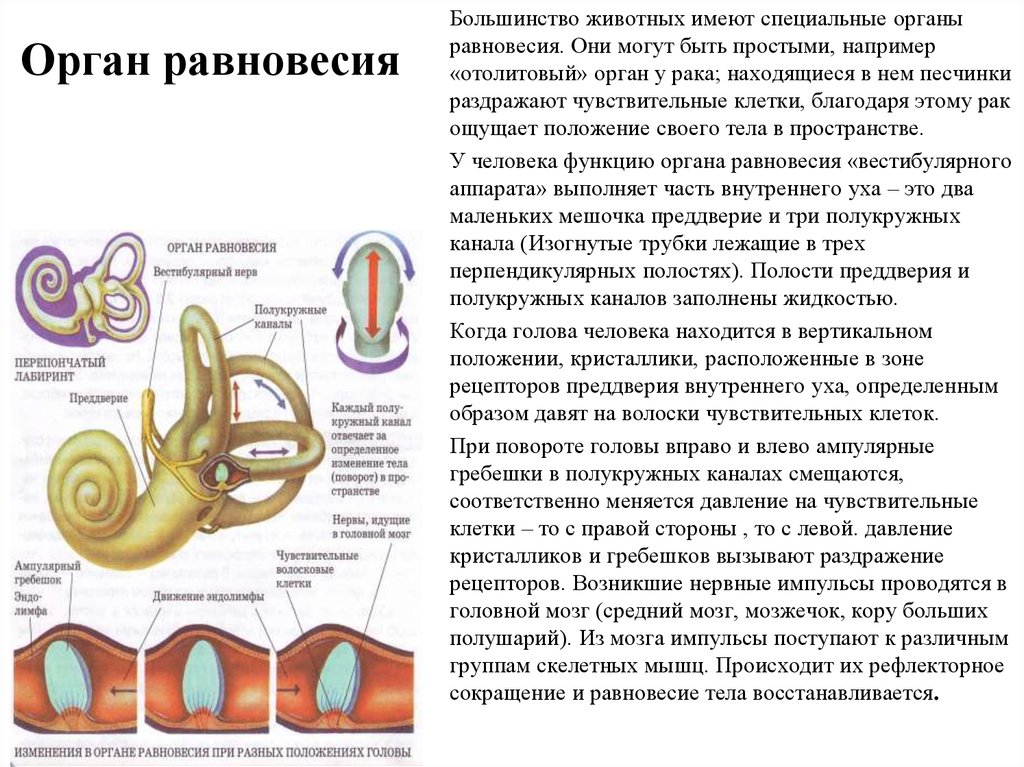 Вверху: На поперечном срезе улитки видны кортиев орган и три лестницы улитки. Внизу: Это показывает структуру кортиева органа, как он появляется в базальном повороте улитки. DC, наружные фаланговые клетки (клетки Дейтерса), поддерживающие наружные волосковые клетки; IPC, внутренние фаланговые клетки, поддерживающие внутреннюю волосковую клетку. (Воспроизведено с разрешения Pickels JO: An Introduction to the Physiology of Hearing, 2nd ed. Academic Press; 1988.)
Вверху: На поперечном срезе улитки видны кортиев орган и три лестницы улитки. Внизу: Это показывает структуру кортиева органа, как он появляется в базальном повороте улитки. DC, наружные фаланговые клетки (клетки Дейтерса), поддерживающие наружные волосковые клетки; IPC, внутренние фаланговые клетки, поддерживающие внутреннюю волосковую клетку. (Воспроизведено с разрешения Pickels JO: An Introduction to the Physiology of Hearing, 2nd ed. Academic Press; 1988.)
улитки и, таким образом, имеет спиралевидную форму. Эта структура содержит узкоспециализированные слуховые рецепторы (волоски) отростки которых пронизывают прочную, мембраноподобную ретикулярной пластинки , которая поддерживается столбчатыми клетками или стержнями Корти (рис. 10-4). Волосковые клетки расположены в четыре ряда: три ряда из наружных волосковых клеток латеральнее туннеля, образованного палочками Корти, и один ряд внутренних волосковых клеток медиально от туннеля. В каждой улитке человека имеется 20 000 наружных волосковых клеток и 3500 внутренних волосковых клеток. Покрывает ряды волосковых клеток тонкая, вязкая, но эластичная текториальная мембрана , в который встроены кончики волосков наружных, но не внутренних волосковых клеток. Тела сенсорных нейронов, разветвляющиеся вокруг оснований волосковых клеток, расположены в спиральном ганглии внутри модиолуса, костного ядра, вокруг которого закручена улитка. От 90 до 95% этих сенсорных нейронов иннервируют внутренние волосковые клетки; только 5–10% иннервируют более многочисленные наружные волосковые клетки, а каждый сенсорный нейрон иннервирует несколько наружных волосковых клеток. Напротив, большинство эфферентных волокон слухового нерва заканчиваются на наружных, а не на внутренних волосковых клетках. Аксоны афферентных нейронов, иннервирующих волосковые клетки, образуют слуховой (улитковый) отдел восьмого черепного нерва.
В каждой улитке человека имеется 20 000 наружных волосковых клеток и 3500 внутренних волосковых клеток. Покрывает ряды волосковых клеток тонкая, вязкая, но эластичная текториальная мембрана , в который встроены кончики волосков наружных, но не внутренних волосковых клеток. Тела сенсорных нейронов, разветвляющиеся вокруг оснований волосковых клеток, расположены в спиральном ганглии внутри модиолуса, костного ядра, вокруг которого закручена улитка. От 90 до 95% этих сенсорных нейронов иннервируют внутренние волосковые клетки; только 5–10% иннервируют более многочисленные наружные волосковые клетки, а каждый сенсорный нейрон иннервирует несколько наружных волосковых клеток. Напротив, большинство эфферентных волокон слухового нерва заканчиваются на наружных, а не на внутренних волосковых клетках. Аксоны афферентных нейронов, иннервирующих волосковые клетки, образуют слуховой (улитковый) отдел восьмого черепного нерва.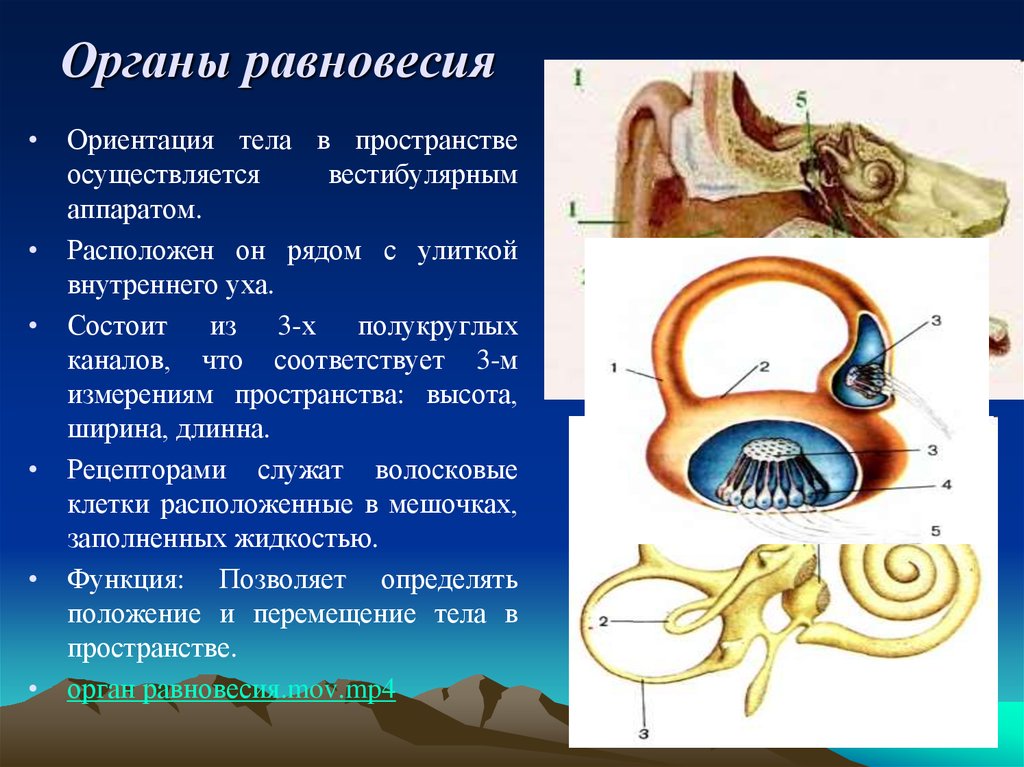
В улитке плотные контакты между волосковыми клетками и соседними фаланговыми клетками не позволяют эндолимфе достигать оснований клеток. Однако базилярная мембрана относительно проницаема для перилимфы в барабанной лестнице, и, следовательно, туннель кортиева органа и основания волосковых клеток омываются перилимфой. Из-за подобных плотных контактов расположение волосковых клеток в других частях внутреннего уха аналогично; то есть отростки волосковых клеток омываются эндолимфой, тогда как их основания омываются перилимфой.
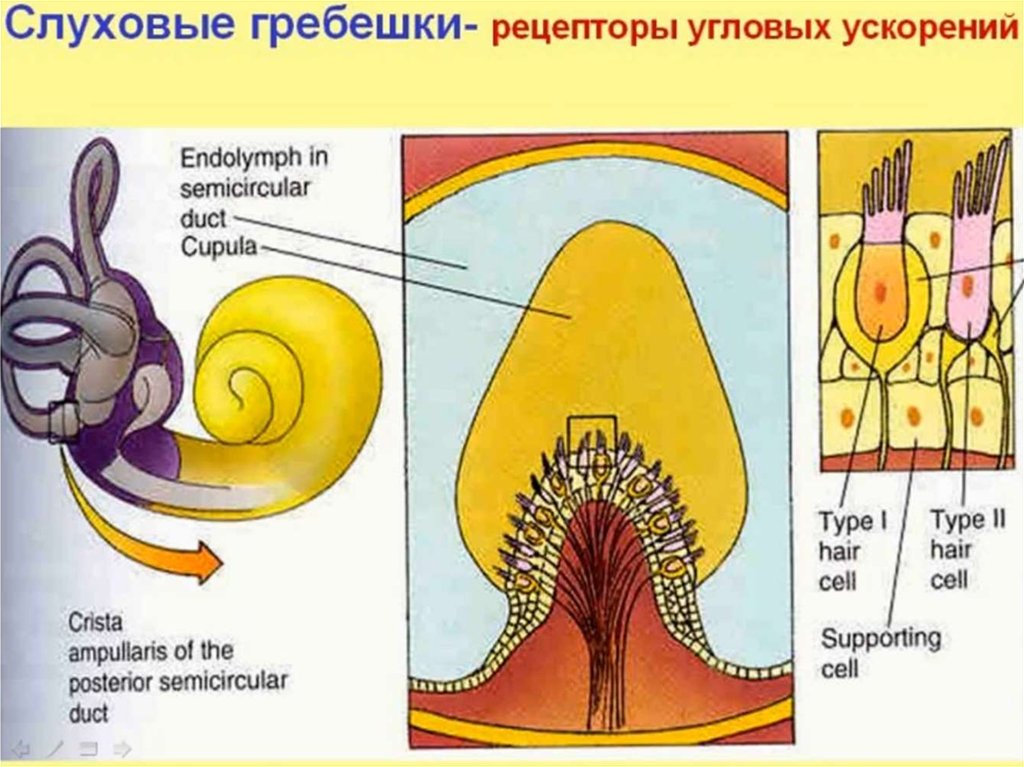
На каждой стороне головы полукружные каналы расположены перпендикулярно друг другу, так что они ориентированы в трех плоскостях пространства. Рецепторная структура, crista ampullaris, , расположена на расширенном конце (ампуле) каждого мембранного канала (рис. 10–3). Каждая криста состоит из волосковых клеток и опорных (поддерживающих) клеток, увенчанных студенистой перегородкой (купулой) , замыкающей ампулу.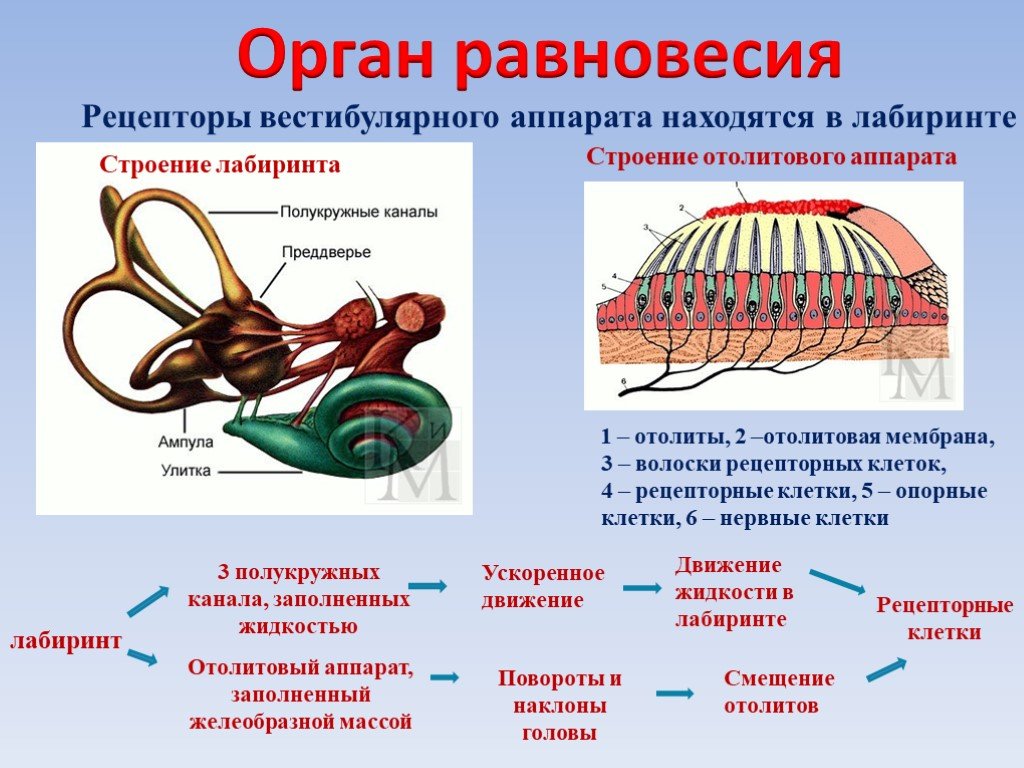 Отростки волосковых клеток заложены в купулу, а основания волосковых клеток находятся в тесном контакте с афферентными волокнами вестибулярный отдел восьмого черепного нерва.
Отростки волосковых клеток заложены в купулу, а основания волосковых клеток находятся в тесном контакте с афферентными волокнами вестибулярный отдел восьмого черепного нерва.
Пара отолитовых органов, мешочек и маточка, расположены вблизи центра перепончатого лабиринта. Чувствующий эпителий этих органов называется макулой . Пятна ориентированы вертикально в мешочке и горизонтально расположены в маточке при вертикальном положении головы. Пятна содержат поддерживающие клетки и волосковые клетки, окруженные отолитовой мембраной, в которую встроены кристаллы карбоната кальция, отолиты (рис. 10–3). Отолиты, которые также называют otoconia или ушной пылью, имеют длину от 3 до 19 мкм у человека. Отростки волосковых клеток встроены в мембрану. Нервные волокна от волосковых клеток соединяются с волокнами от крист в вестибулярном отделе восьмого черепного нерва.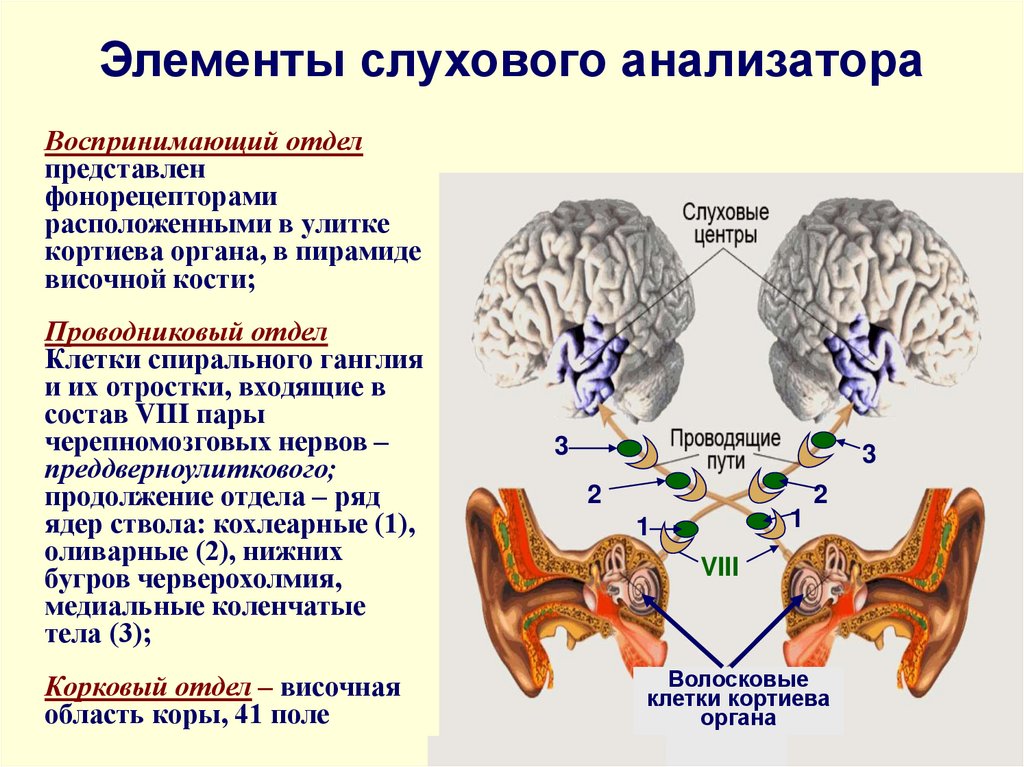
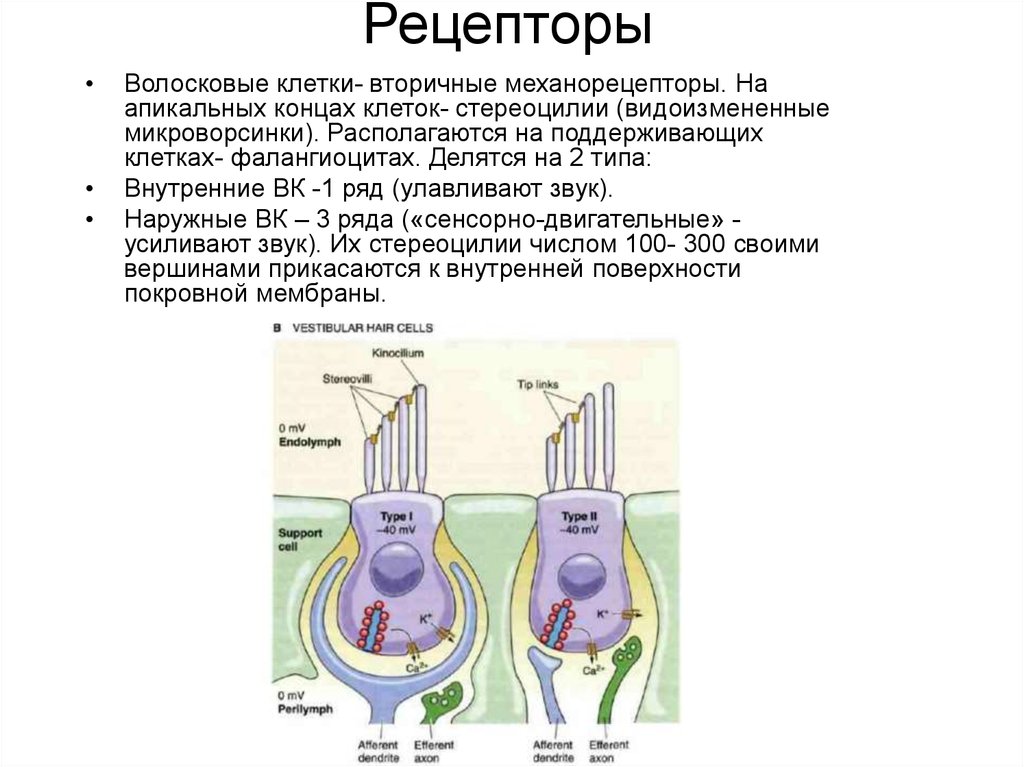
СЕНСОРНЫЕ РЕЦЕПТОРЫ В УХЕ: ВОЛОСКОВЫЕ КЛЕТКИ
Специализированные сенсорные рецепторы уха состоят из шести участков волосковых клеток в перепончатом лабиринте. Это примеры механорецепторы. Волосковые клетки кортиева сигнального органа слуха; волосковые клетки маточки сигнализируют о горизонтальном ускорении; волосковые клетки в мешочке сигнализируют о вертикальном ускорении; и участок в каждом из трех полукружных каналов сигнализирует об ускорении вращения. Эти волосковые клетки имеют общую структуру (рис. 10–5) . Каждый из них встроен в эпителий, состоящий из опорных клеток, базальный конец которого находится в тесном контакте с афферентными нейронами. От апикального конца отходят 30–150 палочковидных отростков, или волосков. За исключением улитки, одна из них, 9Киноцилия 0063, представляет собой настоящую, но неподвижную ресничку с девятью парами микротрубочек по окружности и центральной парой микротрубочек.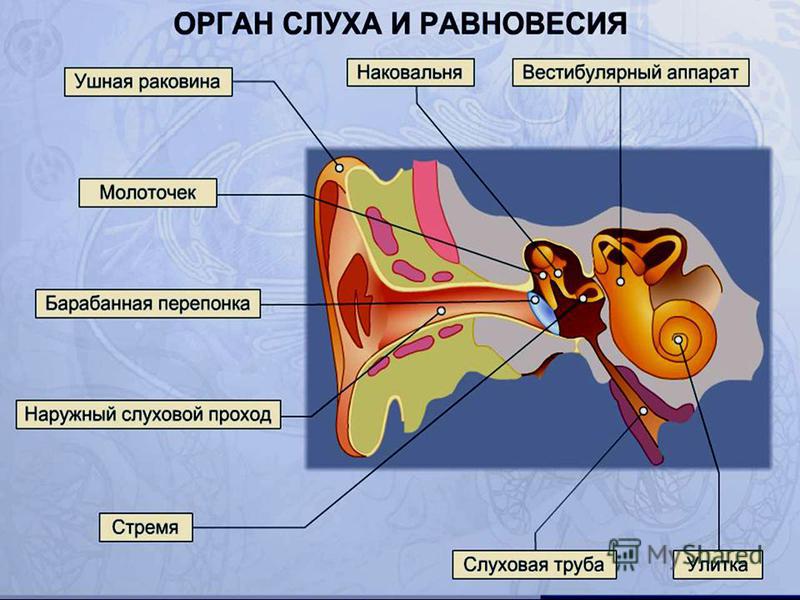 Это один из самых крупных отростков с булавовидным концом. Киноцилия теряется из волосковых клеток улитки у взрослых млекопитающих. Однако другие отростки, называемые стереоцилиями , , присутствуют во всех волосковых клетках. У них есть ядра, состоящие из параллельных нитей актина. Актин покрыт различными изоформами миозина. Внутри скопления отростков на каждой клетке есть упорядоченная структура. По оси к киноцилии стереоцилии прогрессивно увеличиваются в высоту; вдоль перпендикулярной оси все стереоцилии одинаковой высоты.
Это один из самых крупных отростков с булавовидным концом. Киноцилия теряется из волосковых клеток улитки у взрослых млекопитающих. Однако другие отростки, называемые стереоцилиями , , присутствуют во всех волосковых клетках. У них есть ядра, состоящие из параллельных нитей актина. Актин покрыт различными изоформами миозина. Внутри скопления отростков на каждой клетке есть упорядоченная структура. По оси к киноцилии стереоцилии прогрессивно увеличиваются в высоту; вдоль перпендикулярной оси все стереоцилии одинаковой высоты.
РИСУНОК 10–5
Структура волосковой клетки в мешочке. Слева: Волосковые клетки перепончатого лабиринта уха имеют общую структуру, и каждая из них находится внутри эпителия опорных клеток (SC), увенчанных отолитовой мембраной (OM), залитой кристаллами карбоната кальция, отолитами (OL). От апикального конца отходят палочковидные отростки, или волосковые клетки (ВК), контактирующие с афферентными (А) и эфферентными (Е) нервными волокнами.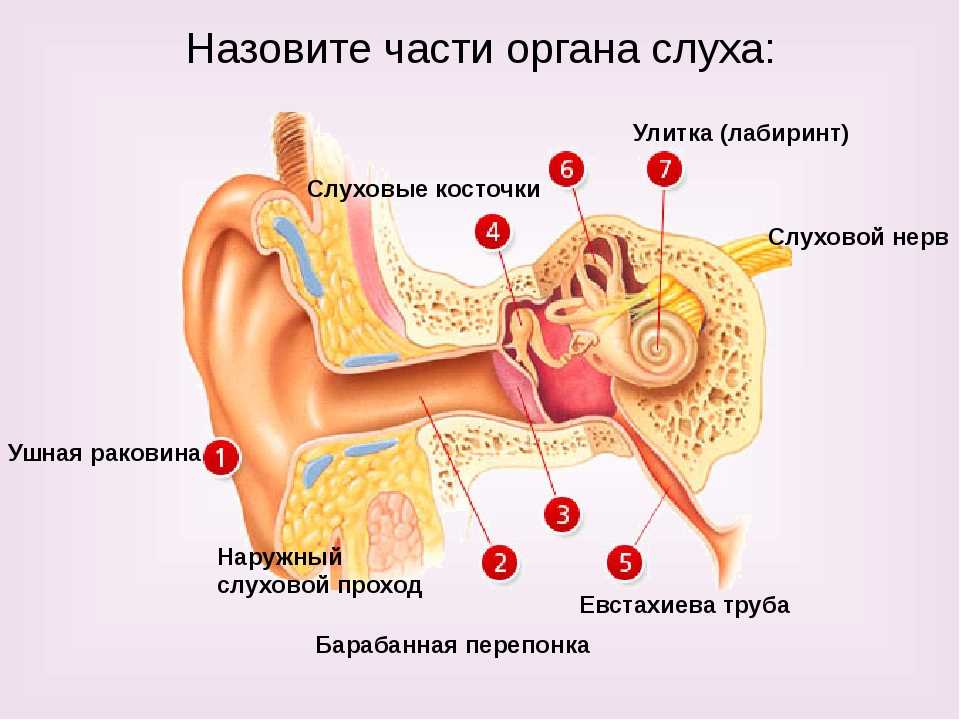 За исключением улитки, одна из них, 9Киноцилия 0063 (K) представляет собой настоящую, но неподвижную ресничку с девятью парами микротрубочек по окружности и центральной парой микротрубочек. Другие отростки, стереоцилии (S), обнаруживаются во всех волосковых клетках; у них есть ядра актиновых филаментов, покрытых изоформами миозина. Внутри скопления отростков на каждой клетке есть упорядоченная структура. По оси к киноцилии стереоцилии прогрессивно увеличиваются в высоту; вдоль перпендикулярной оси все стереоцилии имеют одинаковую высоту. (Воспроизведено с разрешения Llinas R, Precht W (eds): Нейробиология лягушки . Спрингер; 1976.) Справа: Сканирующая электронная микрофотография отростков волосковой клетки в мешочке. Отолитовая мембрана удалена. Небольшие выступы вокруг волосковой клетки представляют собой микроворсинки на опорных клетках. (Используется с разрешения AJ Hudspeth.)
За исключением улитки, одна из них, 9Киноцилия 0063 (K) представляет собой настоящую, но неподвижную ресничку с девятью парами микротрубочек по окружности и центральной парой микротрубочек. Другие отростки, стереоцилии (S), обнаруживаются во всех волосковых клетках; у них есть ядра актиновых филаментов, покрытых изоформами миозина. Внутри скопления отростков на каждой клетке есть упорядоченная структура. По оси к киноцилии стереоцилии прогрессивно увеличиваются в высоту; вдоль перпендикулярной оси все стереоцилии имеют одинаковую высоту. (Воспроизведено с разрешения Llinas R, Precht W (eds): Нейробиология лягушки . Спрингер; 1976.) Справа: Сканирующая электронная микрофотография отростков волосковой клетки в мешочке. Отолитовая мембрана удалена. Небольшие выступы вокруг волосковой клетки представляют собой микроворсинки на опорных клетках. (Используется с разрешения AJ Hudspeth.)
ЭЛЕКТРИЧЕСКИЕ РЕАКЦИИ
Очень тонкие процессы, называемые каждый стереоцилий в сторону своего более высокого соседа, а на стыке находятся механически чувствительные катионные каналы в более высоком процессе.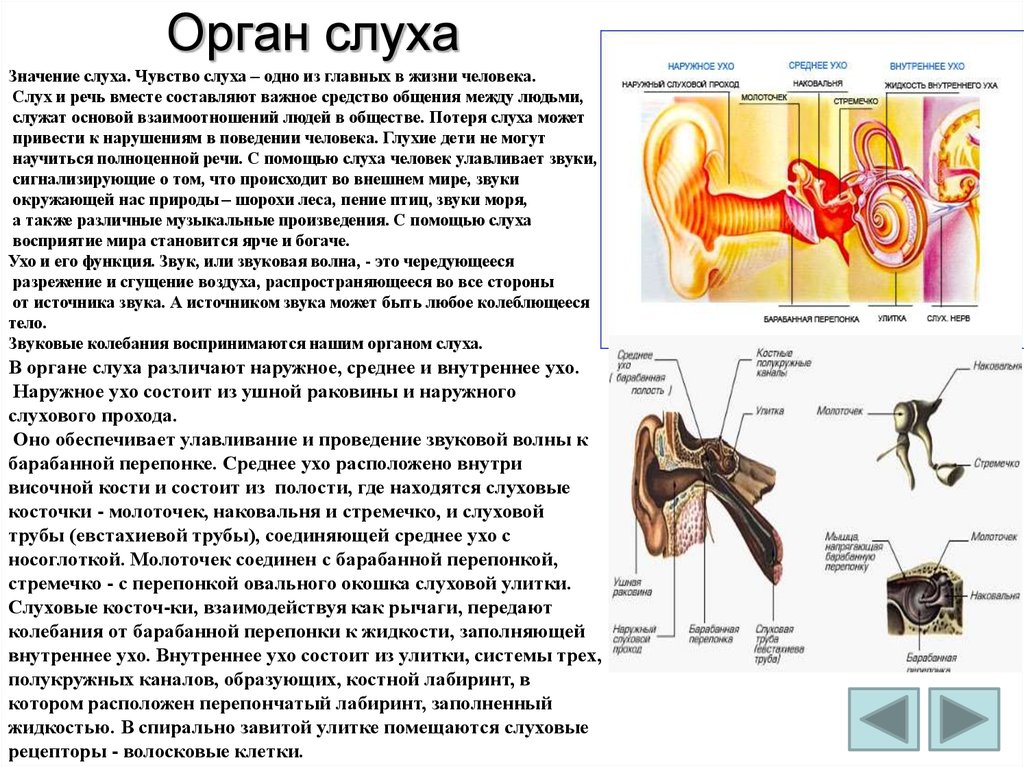 Когда более короткие стереоцилии смещаются к более высоким, время открытия этих каналов увеличивается. К + , наиболее распространенный катион в эндолимфе, и Ca 2+ проникают через канал и вызывают деполяризацию. Молекулярный двигатель на основе миозина в более высоком соседе затем перемещает канал к основанию, ослабляя напряжение в концевом звене (рис. 10-6). Это приводит к закрытию канала и позволяет восстановить состояние покоя. Деполяризация волосковых клеток заставляет их высвобождать нейротрансмиттер, вероятно, глутамат, который инициирует деполяризацию соседних афферентных нейронов.
Когда более короткие стереоцилии смещаются к более высоким, время открытия этих каналов увеличивается. К + , наиболее распространенный катион в эндолимфе, и Ca 2+ проникают через канал и вызывают деполяризацию. Молекулярный двигатель на основе миозина в более высоком соседе затем перемещает канал к основанию, ослабляя напряжение в концевом звене (рис. 10-6). Это приводит к закрытию канала и позволяет восстановить состояние покоя. Деполяризация волосковых клеток заставляет их высвобождать нейротрансмиттер, вероятно, глутамат, который инициирует деполяризацию соседних афферентных нейронов.
РИСУНОК 10–6
Схематическое представление роли концевых связей в реакциях волосковых клеток. Когда стереоцилию подталкивают к более высокой стереоцилии, кончик звена растягивается и открывает ионный канал в своем более высоком соседе. Затем канал, по-видимому, перемещается вниз по более высокой стереоцилии с помощью молекулярного мотора, поэтому напряжение на концевом звене ослабевает.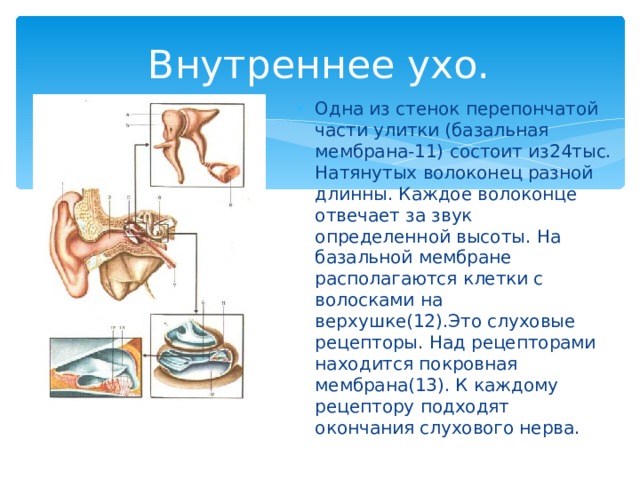 Когда волоски возвращаются в исходное положение, двигатель движется обратно вверх по стереоцилии.
Когда волоски возвращаются в исходное положение, двигатель движется обратно вверх по стереоцилии.
K + , который проникает в волосковые клетки через механически чувствительные катионные каналы, рециркулируется (рис. 10–7) . Он проникает в опорные клетки и затем переходит к другим опорным клеткам посредством плотных контактов. В улитке он в конечном итоге достигает сосудистой полоски и секретируется обратно в эндолимфу, завершая цикл.
РИСУНОК 10–7
Ионный состав перилимфы вестибулярной лестницы, эндолимфы средней лестницы и перилимфы барабанной лестницы. SL, спиральная связка. СВ, сосудистая полоска. Пунктирная стрелка указывает путь, по которому K + рециркулирует из волосковых клеток в поддерживающие клетки и спиральную связку, а затем секретируется обратно в эндолимфу клетками сосудистой полоски.
Как описано выше, отростки волосковых клеток проецируются в эндолимфу, тогда как основания омываются перилимфой. Такое расположение необходимо для нормального производства рецепторных потенциалов. Перилимфа образуется в основном из плазмы. С другой стороны, эндолимфа образуется в средней лестнице сосудистой полоской и имеет высокую концентрацию К9.0134 + и низкой концентрации Na + (рис. 10–7). Клетки сосудистой полоски имеют высокую концентрацию Na + , K + АТФазы. Кроме того, оказалось, что уникальная электрогенная помпа K + в сосудистой полоске объясняет тот факт, что средняя лестница электрически положительна на 85 мВ относительно вестибулярной лестницы и барабанной лестницы.
Такое расположение необходимо для нормального производства рецепторных потенциалов. Перилимфа образуется в основном из плазмы. С другой стороны, эндолимфа образуется в средней лестнице сосудистой полоской и имеет высокую концентрацию К9.0134 + и низкой концентрации Na + (рис. 10–7). Клетки сосудистой полоски имеют высокую концентрацию Na + , K + АТФазы. Кроме того, оказалось, что уникальная электрогенная помпа K + в сосудистой полоске объясняет тот факт, что средняя лестница электрически положительна на 85 мВ относительно вестибулярной лестницы и барабанной лестницы.
Мембранный потенциал покоя волосковых клеток составляет около –60 мВ. Когда стереоцилии подталкиваются к киноцилии, мембранный потенциал снижается примерно до -50 мВ. Когда пучок отростков смещается в противоположном направлении, клетка гиперполяризуется. Смещение отростков в направлении, перпендикулярном этой оси, не приводит к изменению мембранного потенциала, а смещение отростков в направлениях, промежуточных между этими двумя направлениями, вызывает деполяризацию или гиперполяризацию, пропорциональную степени, в которой направление направлено к мембране или от нее.